
Miért létfontosságú a sejtek számára, hogy folyamatosan anyagokat cseréljenek környezetükkel, és hogyan biztosítják ezt a precíz, mégis rendkívül komplex folyamatot? A válasz a transzporttal rendelkező cella mélyreható működésében rejlik, amely nem csupán a túlélés alapja, hanem az élet szinte minden biológiai funkciójának mozgatórugója is. A sejt, mint az élet alapegysége, egy dinamikus rendszer, amely állandó interakcióban áll környezetével, és ezen interakciók központi eleme az anyagok szervezett áramlása a membránokon keresztül.
Ahhoz, hogy megértsük a transzporttal rendelkező cella jelentőségét és működését, először is a sejtmembrán szerkezetét kell alaposan megvizsgálnunk. Ez a hajszálvékony, kettős lipidréteg nem csupán egy fizikai határ, hanem egy rendkívül szelektív és intelligens kapu, amely szabályozza a be- és kiáramló molekulák útját. A membrán áteresztőképessége, vagyis szelektív permeabilitása, teszi lehetővé, hogy a sejt fenntartsa belső környezetének stabilitását, a homeosztázist, ami elengedhetetlen a megfelelő működéshez és a túléléshez.
A sejtmembrán, mint a transzport központja
A plazmamembrán, vagy sejtmembrán, egy lenyűgöző szerkezet, amely alapvetően egy foszfolipid kettős rétegből épül fel. Ezek a foszfolipid molekulák egy hidrofób (víztaszító) belső részt és egy hidrofil (vízkedvelő) külső részt alkotnak, ami meghatározza a membrán alapvető áteresztőképességi tulajdonságait. A kisebb, zsíroldékony molekulák, mint például az oxigén, a szén-dioxid vagy a szteroid hormonok, viszonylag könnyedén átjuthatnak ezen a lipidrétegen.
Azonban a nagyobb, töltött vagy vízkedvelő molekulák, mint az ionok, a glükóz vagy az aminosavak, nem képesek egyszerűen átdiffundálni a hidrofób belső részen. Itt lépnek képbe a membránfehérjék, amelyek beágyazódnak a lipid kettős rétegbe, vagy ahhoz kapcsolódnak. Ezek a fehérjék a transzportfolyamatok döntő szereplői, hiszen ők biztosítják a specifikus és szabályozott átjutást a sejt belsejébe és onnan kifelé.
A sejtmembrán nem csupán egy fizikai határ, hanem egy dinamikus és intelligens felület, amely aktívan szabályozza a sejt és környezete közötti anyagcserét.
A membránfehérjék rendkívül változatosak funkciójukat tekintve. Vannak köztük integráns membránfehérjék, amelyek teljesen átérnek a kettős lipidrétegen, és perifériás membránfehérjék, amelyek a membrán felületéhez kapcsolódnak. A transzportfolyamatok szempontjából az integráns fehérjék, különösen a transzmembrán fehérjék, a legfontosabbak, hiszen ők alkotják azokat a csatornákat, hordozókat és pumpákat, amelyek az anyagok mozgását irányítják.
A sejtmembrán fluid mozaik modellje szerint ezek a fehérjék nem rögzítettek, hanem viszonylag szabadon mozognak a lipid kettős rétegen belül. Ez a fluiditás elengedhetetlen a membrán funkcióinak, így a transzportfolyamatoknak is a hatékony végrehajtásához. A membrán glikolipidekkel és glikoproteinekkel is gazdagított, amelyek a sejt felismerésében és a jelátvitelben is szerepet játszanak, közvetetten befolyásolva a transzport szabályozását.
A transzport alapelvei: gradiens és energia
A sejten belüli és kívüli anyagmozgást alapvetően két fő tényező, a koncentrációgradiens és az elektromos gradiens határozza meg. Ezek együttesen alkotják az elektrokémiai gradienst, amely a legtöbb ion transzportjának hajtóereje. A koncentrációgradiens egyszerűen azt jelenti, hogy egy adott anyagból eltérő mennyiség van a membrán két oldalán, és az anyag hajlamos a magasabb koncentrációjú helyről az alacsonyabb felé mozogni.
Az elektromos gradiens a töltött részecskék, az ionok mozgását befolyásolja. A sejtmembránon belül és kívül eltérő az elektromos töltés eloszlása, ami egy membránpotenciált eredményez. A legtöbb sejt belseje negatívabb töltésű a külsejéhez képest. Ez az elektromos különbség vonzza a pozitív ionokat a sejt belsejébe, és taszítja a negatív ionokat.
A transzportfolyamatok energiaigény szempontjából két fő kategóriába sorolhatók: passzív transzportra és aktív transzportra. A passzív transzport nem igényel közvetlen metabolikus energiát, mivel az anyagok az elektrokémiai gradiensük mentén mozognak. Ezzel szemben az aktív transzport energiát igényel, általában ATP hidrolízis formájában, hogy az anyagokat a gradiensük ellenében mozgassa.
Passzív transzport: a gradiens mentén
A passzív transzport a sejtek legkevésbé energiaigényes szállítási mechanizmusa, mivel a molekulák a meglévő elektrokémiai potenciálkülönbségüket kihasználva jutnak át a membránon. Ennek több formája is létezik, amelyek a molekulák méretétől, töltésétől és a membránnal való kölcsönhatásától függően különböznek.
Egyszerű diffúzió
Az egyszerű diffúzió a passzív transzport legegyszerűbb formája, amely során a molekulák közvetlenül átjutnak a lipid kettős rétegen. Ez a mechanizmus a kis, apoláris, zsíroldékony molekulákra jellemző, mint például az oxigén (O2), a szén-dioxid (CO2), a nitrogén (N2), a benzol vagy az etanol. Ezek a molekulák a magasabb koncentrációjú területről az alacsonyabb koncentrációjú területre vándorolnak, amíg egyensúly nem alakul ki a membrán két oldalán.
Az egyszerű diffúzió sebessége függ a molekula méretétől, zsíroldékonyságától és a koncentrációkülönbség nagyságától. Minél kisebb és zsíroldékonyabb egy molekula, és minél nagyobb a koncentrációgradiens, annál gyorsabban diffundál. Ez a folyamat nem telíthető, ami azt jelenti, hogy a transzport sebessége lineárisan arányos a koncentrációkülönbséggel, ellentétben a hordozófehérjékkel történő transzporttal.
Facilitált diffúzió
A facilitált diffúzió szintén passzív folyamat, ami azt jelenti, hogy nem igényel közvetlen energiabefektetést, és az anyagok az elektrokémiai gradiensük mentén mozognak. A különbség az egyszerű diffúzióhoz képest az, hogy ebben az esetben speciális membránfehérjék, úgynevezett hordozófehérjék vagy csatornafehérjék segítik az anyagok átjutását. Ezek a fehérjék specifikusak, azaz csak bizonyos molekulákat képesek szállítani.
Csatornafehérjék és ioncsatornák
A csatornafehérjék hidrofób pórusokat vagy csatornákat képeznek a membránon keresztül, amelyeken keresztül a megfelelő méretű és töltésű ionok vagy kis molekulák gyorsan áthaladhatnak. A legfontosabb csatornafehérjék az ioncsatornák, amelyek specifikusak bizonyos ionok, például Na+, K+, Ca2+ vagy Cl- számára. Ezek az ioncsatornák gyakran szabályozottak, ami azt jelenti, hogy nyitott vagy zárt állapotban lehetnek, és különböző ingerekre reagálnak.
Az ioncsatornák szabályozásának módjai a következők:
- Feszültségfüggő csatornák: Ezek a csatornák a membránpotenciál változására nyílnak vagy záródnak. Kritikusak az idegsejtek és izomsejtek működésében, például az akciós potenciál kialakításában.
- Ligandfüggő csatornák (receptorok): Ezek a csatornák egy specifikus molekula (ligand), például egy neurotranszmitter vagy hormon kötődésére nyílnak meg. Példák erre az acetilkolin receptorok a neuromuszkuláris junkcióban.
- Mechanikusan kapuzott csatornák: Fizikai ingerekre, mint például nyomásra vagy feszülésre nyílnak. Fontosak a tapintás, a hallás és az egyensúly érzékelésében.
Az aquaporinok egy speciális típusú csatornafehérjék, amelyek a vízmolekulák rendkívül gyors átjutását teszik lehetővé a membránon keresztül. Bár a víz képes lassan átdiffundálni a lipid kettős rétegen is (ozmózis), az aquaporinok drámaian felgyorsítják ezt a folyamatot, ami létfontosságú például a vesékben a vízvisszaszívás szempontjából.
Hordozófehérjék (transzporterek)
A hordozófehérjék más mechanizmussal működnek, mint a csatornák. Ezek a fehérjék először megkötik a szállítandó molekulát a membrán egyik oldalán, majd konformációs változáson mennek keresztül, ami lehetővé teszi a molekula átjutását a membrán másik oldalára. A hordozófehérjék lassabban szállítanak, mint a csatornák, és telíthetőek, ami azt jelenti, hogy van egy maximális szállítási sebességük, amit a hordozófehérjék száma és működési sebessége korlátoz.
Jó példa a hordozófehérjékre a glükóz transzporterek (GLUT) családja. Ezek a fehérjék felelősek a glükóz felvételéért a vérből a sejtekbe, például az izom- és zsírsejtekbe (GLUT4), vagy a vörösvértestekbe (GLUT1). A GLUT4 transzporter inzulinra érzékeny, ami kulcsszerepet játszik a vércukorszint szabályozásában.
A facilitált diffúzió lehetővé teszi a specifikus és szabályozott anyagfelvételt a sejt számára anélkül, hogy energiát kellene fektetnie a folyamatba.
Ozmózis
Az ozmózis a vízmolekulák diffúziója egy féligáteresztő membránon keresztül, a magasabb vízkoncentrációjú területről (azaz alacsonyabb oldott anyag koncentrációjú, hipotóniás oldatból) az alacsonyabb vízkoncentrációjú területre (azaz magasabb oldott anyag koncentrációjú, hipertóniás oldatba). Ez a passzív folyamat alapvető fontosságú a sejtek térfogatának és belső nyomásának fenntartásában.
Ha egy sejtet izotóniás oldatba helyezünk, ahol az oldott anyagok koncentrációja megegyezik a sejt belsejével, akkor nincs nettó vízáramlás. Hipotóniás oldatban a víz beáramlik a sejtbe, ami duzzadáshoz, sőt akár szétrepedéshez (lízihez) vezethet, különösen állati sejtekben. Hipertóniás oldatban a víz kiáramlik a sejtből, ami a sejt zsugorodásához (plasmolysis) vezet.
Növényi sejtekben a sejtfal megakadályozza a sejt szétrepedését hipotóniás környezetben, és segít fenntartani a turgornyomást, ami a növényi szövetek merevségét adja. Az ozmózis tehát alapvető szerepet játszik a sejtek vízháztartásában és az egész szervezet folyadékegyensúlyában.
Aktív transzport: a gradiens ellenében
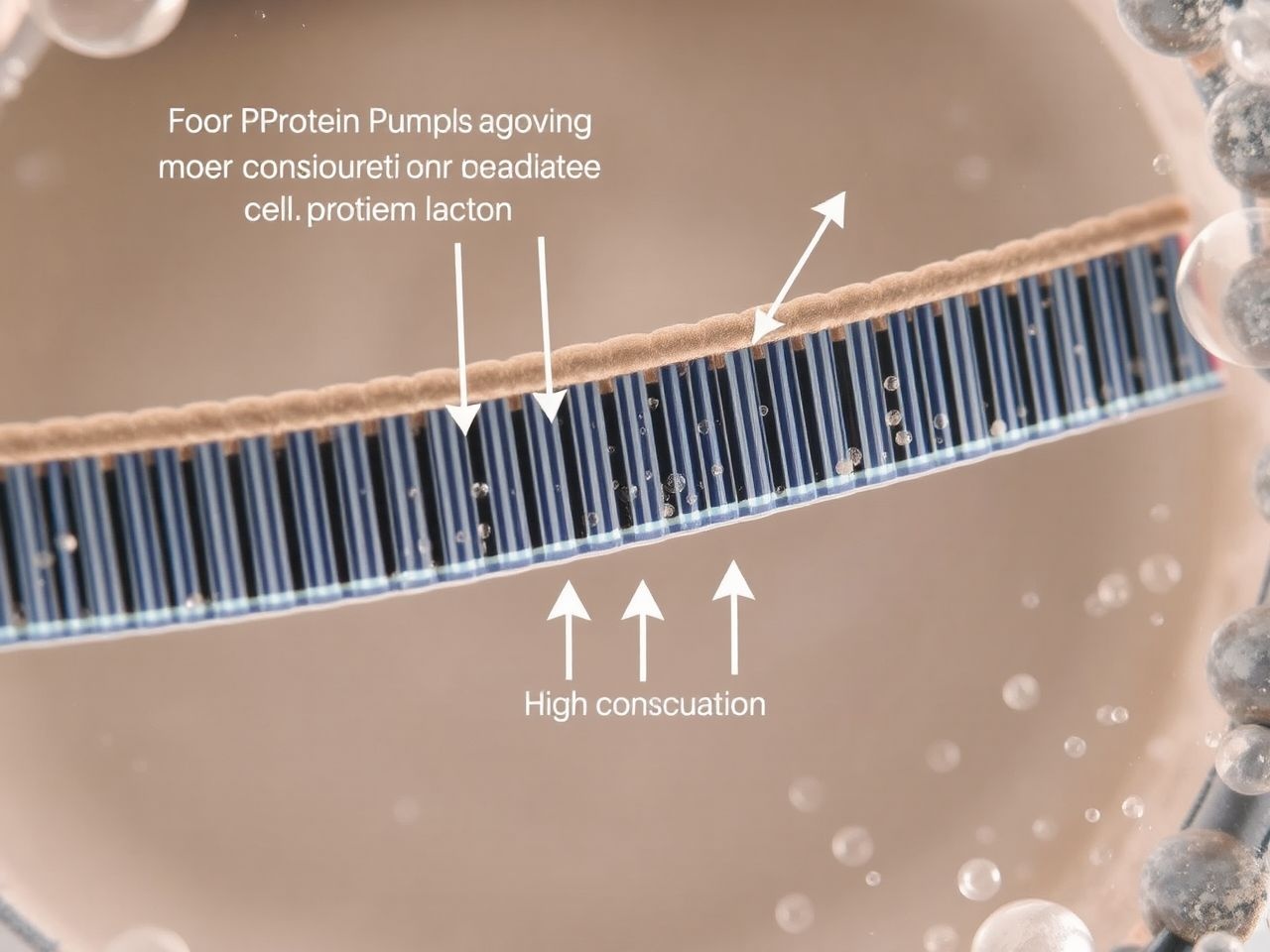
Az aktív transzport olyan folyamat, amely során a sejtek energiát használnak fel az anyagok mozgatására az elektrokémiai gradiensük ellenében, azaz az alacsonyabb koncentrációjú területről a magasabb koncentrációjú területre. Ez a mechanizmus elengedhetetlen ahhoz, hogy a sejt fenntartsa a belső környezetét, és felvegye a számára szükséges anyagokat, még akkor is, ha azok koncentrációja alacsonyabb a külvilágban.
Két fő típusa van az aktív transzportnak: a primer aktív transzport és a szekunder aktív transzport.
Primer aktív transzport
A primer aktív transzport során a transzporter fehérje közvetlenül használja fel az ATP hidrolíziséből származó energiát a molekulák szállítására. Ezeket a transzportereket gyakran pumpáknak nevezzük. Az ATP hidrolízise során felszabaduló energia konformációs változást okoz a fehérjében, ami lehetővé teszi az ionok vagy molekulák mozgatását a membránon keresztül.
Na+/K+-ATPáz pumpa (nátrium-kálium pumpa)
A Na+/K+-ATPáz pumpa az egyik legfontosabb primer aktív transzporter az állati sejtekben, és a sejt energiafelhasználásának jelentős részéért felelős. Ez a pumpa a nátrium-ionokat (Na+) pumpálja ki a sejtből, és a kálium-ionokat (K+) pumpálja be a sejtbe, mindkettőt az elektrokémiai gradiensük ellenében. Minden egyes ciklusban 3 Na+ iont juttat ki a sejtből, és 2 K+ iont juttat be a sejtbe, egyetlen ATP molekula hidrolízisének felhasználásával.
Ennek a pumpának több létfontosságú funkciója is van:
- Membránpotenciál fenntartása: A pumpa által létrehozott töltéskülönbség hozzájárul a negatív nyugalmi membránpotenciál fenntartásához, ami alapvető az ideg- és izomsejtek ingerlékenységéhez.
- Ozmotikus egyensúly fenntartása: A Na+ ionok aktív kipumpálása megakadályozza a víz beáramlását és a sejt duzzadását, ezzel fenntartva a sejt térfogatát.
- Szekunder aktív transzport hajtóereje: A sejt belsejében kialakított alacsony Na+ koncentrációgradiens energiát biztosít más, szekunder aktív transzporterek számára.
A Na+/K+-ATPáz működésének részletesebb megértése kulcsfontosságú számos élettani folyamat, például az idegi impulzusok terjedése vagy a vesék működése szempontjából. Gyógyszerek, mint például a digoxin, a szívizomsejtek Na+/K+-ATPáz pumpáját gátolva fejtik ki hatásukat, erősítve a szív összehúzódását.
Ca2+-ATPáz pumpa (kalcium pumpa)
A Ca2+-ATPáz pumpák felelősek a kalciumionok (Ca2+) aktív kiszállításáért a citoplazmából, vagy a szarkoplazmatikus retikulum (izomsejtekben) vagy az endoplazmatikus retikulum (más sejtekben) lumenébe. A citoplazmatikus Ca2+ koncentráció rendkívül alacsonyan tartása létfontosságú, mivel a Ca2+ számos sejten belüli jelátviteli útvonal kulcsfontosságú másodlagos hírvivője.
Az izomösszehúzódás során például a Ca2+ felszabadul a szarkoplazmatikus retikulumból, majd a kontrakció befejezése után a Ca2+-ATPáz pumpák aktívan visszapumpálják oda, relaxációt eredményezve. A pumpa hibás működése számos betegséghez, többek között izom- és szívbetegségekhez is vezethet.
H+-ATPáz pumpa (proton pumpa)
A H+-ATPáz pumpák, vagy proton pumpák, hidrogénionokat (H+) pumpálnak a membránon keresztül, szintén ATP hidrolízis felhasználásával. Ezek a pumpák különösen fontosak a savas környezet fenntartásában, például a gyomorban (gyomor savtermelése), a lizoszómákban (enzimek optimális működéséhez) és a vesékben (vizelet savanyítása).
Az aktív transzport biztosítja a sejt számára a képességet, hogy a külső környezettel szemben is fenntartsa a belső egyensúlyát, és felhalmozza a létfontosságú anyagokat.
A gyomorsav túlzott termelése esetén a proton pumpa gátlók, mint például az omeprazol, hatékony gyógyszerek, amelyek csökkentik a gyomorsav kiválasztását a gyomor parietális sejtjeiben.
Szekunder aktív transzport (ko-transzport)
A szekunder aktív transzport során az anyagok mozgatása a gradiensük ellenében történik, de nem közvetlenül ATP hidrolízisből nyert energiát használ. Ehelyett egy másik ion (általában Na+ vagy H+) elektrokémiai gradiensében tárolt energiát hasznosítja, amelyet egy primer aktív transzporter hozott létre. Ez a folyamat két molekula egyidejű szállítását foglalja magában: az egyik a gradiensével megegyező irányban mozog, a másik a gradiensével ellentétes irányban.
Két fő típusa van a szekunder aktív transzportnak:
- Szimporterek (ko-transzporterek): A két molekula azonos irányba mozog a membránon keresztül.
- Antiporterek (ellen-transzporterek): A két molekula ellentétes irányba mozog a membránon keresztül.
Szimporterek (Na+/glükóz ko-transzporter, SGLT)
A szimporterek példája a vékonybél hámsejtjeiben és a vesetubulusokban található Na+/glükóz ko-transzporter (SGLT). Ez a transzporter a nátriumionok befelé irányuló, a gradienssel megegyező mozgását használja fel arra, hogy a glükózt a gradiensével ellentétesen, a sejtbe juttassa. Mivel a Na+ koncentrációja a sejt belsejében alacsony a Na+/K+-ATPáz működésének köszönhetően, a Na+ befelé áramlása „húzza magával” a glükózt.
Az SGLT1 és SGLT2 transzporterek kulcsszerepet játszanak a táplálékból származó glükóz felszívódásában és a vesékben a glükóz visszaszívásában. Az SGLT2 gátlók új gyógyszercsoportot képviselnek a 2-es típusú cukorbetegség kezelésében, mivel fokozzák a glükóz vizelettel történő kiválasztását.
Antiporterek (Na+/Ca2+ kicserélő, NCX)
Az antiporterek, mint például a Na+/Ca2+ kicserélő (NCX), a membránon keresztül ellentétes irányban mozgatnak két különböző iont. Az NCX a nátriumionok befelé irányuló áramlását használja fel arra, hogy a kalciumionokat kipumpálja a sejtből, a gradiensük ellenében. Ez a mechanizmus különösen fontos a szívizomsejtekben a Ca2+ koncentráció szabályozásában, hozzájárulva a relaxációhoz az összehúzódás után.
Egy másik példa az antiporterekre a Cl-/HCO3- kicserélő, amely a vörösvértestekben a szén-dioxid szállításában játszik szerepet azáltal, hogy kloridionokat cserél bikarbonátionokra.
Vezikuláris transzport: nagy molekulák és sejtrészek szállítása
A kis molekulák és ionok membránon keresztüli transzportja mellett a sejteknek szükségük van nagyobb molekulák, sőt akár egész sejtrészek vagy mikroorganizmusok szállítására is. Erre szolgál a vezikuláris transzport, amely energiát igénylő folyamat, és membránnal határolt hólyagocskák, úgynevezett vezikulák segítségével zajlik. A vezikuláris transzport két fő típusa az endocitózis (anyagok felvétele) és az exocitózis (anyagok kiválasztása).
Endocitózis: anyagok felvétele a sejtbe
Az endocitózis során a sejtmembrán befűződik, körülveszi a felvenni kívánt anyagot, majd egy vezikulát képezve bejuttatja azt a sejt belsejébe. Három fő típusa van:
Fagocitózis (sejtevés)
A fagocitózis a „sejtevés” folyamata, amely során a sejt nagy részecskéket, például baktériumokat, sejttörmeléket vagy elhalt sejteket kebelez be. Ez a folyamat specializált sejtekre, például a makrofágokra és a neutrofilekre (fehérvérsejtek) jellemző, amelyek az immunrendszer fontos részei. A bekebelezett részecske egy nagy vezikulába, az úgynevezett fagoszómába kerül, amely ezután fúzionál a lizoszómákkal, ahol az emésztőenzimek lebontják a tartalmat.
Pinocitózis (sejtivás)
A pinocitózis a „sejtivás” folyamata, amely során a sejt folyékony anyagokat vagy oldott makromolekulákat vesz fel. A sejtmembrán apró vezikulákat képezve bekebelezi a külső folyadékcseppeket a benne oldott anyagokkal együtt. Ez egy nem specifikus folyamat, mivel a sejt egyszerűen mintát vesz a környező folyadékból. Minden eukarióta sejt képes pinocitózisra, és ez a mechanizmus fontos a sejt táplálékfelvételében és a sejtfelszín újrahasznosításában.
Receptor-mediált endocitózis
A receptor-mediált endocitózis egy rendkívül specifikus és hatékony mechanizmus, amely lehetővé teszi a sejt számára, hogy nagy mennyiségben vegyen fel specifikus molekulákat a környezetéből. Ebben a folyamatban a felvenni kívánt molekulák (ligandok) először a sejt felszínén lévő specifikus receptorokhoz kötődnek. Ezek a receptorok egy membránterületen koncentrálódnak, amelyet gyakran klatrinnal borított gödörnek (clathrin-coated pit) neveznek.
A ligand-receptor komplexek kötődése kiváltja a klatrin bevonatú gödör befűződését és egy klatrinnal borított vezikula képződését. Ez a vezikula ezután a sejt belsejébe vándorol, ahol a klatrin bevonat leválik, és a vezikula fúzionál egy endoszómával. Az endoszómában a receptorok általában leválnak a ligandokról, és visszatérnek a sejt felszínére, míg a ligandokat tartalmazó vezikulák tovább jutnak a lizoszómákba lebontásra vagy a sejten belüli felhasználásra.
Ez a mechanizmus létfontosságú például a koleszterin felvételében LDL-partikulák formájában, vas felvételében transzferrin segítségével, vagy a hormonok és növekedési faktorok sejtekbe jutásában. A receptor-mediált endocitózis zavara súlyos betegségekhez vezethet, mint például a familiáris hiperkoleszterinémia.
Exocitózis: anyagok kiválasztása a sejtből
Az exocitózis az a folyamat, amely során a sejt a belsejében szintetizált anyagokat vagy a lebontásra szánt salakanyagokat juttatja ki a sejtből. Ez is vezikulák segítségével történik. A sejt belsejében, például a Golgi-készülékben képződő vezikulák a plazmamembránhoz vándorolnak, fúzionálnak vele, és tartalmukat a sejten kívüli térbe ürítik.
Az exocitózisnak két fő útja van:
- Konstitutív exocitózis: Ez egy folyamatosan zajló folyamat, amely minden sejtben megfigyelhető. Fontos a membránfehérjék és -lipidek beépítésében a plazmamembránba, valamint a sejten kívüli térbe ürítendő anyagok, például a kollagén vagy az immunglobulinok folyamatos kiválasztásában.
- Szabályozott exocitózis: Ez a folyamat csak specifikus ingerek hatására aktiválódik, és a sejt által termelt, raktározott anyagok, például hormonok (inzulin), neurotranszmitterek (acetilkolin) vagy emésztőenzimek (hasnyálmirigy enzimjei) gyors és kontrollált felszabadítására szolgál. A vezikulák ezekben az esetekben a sejtmembrán közelében várakoznak, és csak egy jel (pl. Ca2+ beáramlás) hatására fúzionálnak a membránnal.
Az exocitózis alapvető fontosságú az idegsejtek közötti kommunikációban (szinaptikus transzmisszió), a hormontermelő sejtek működésében és a sejtfal képződésében a növényekben. A SNARE fehérjék kulcsszerepet játszanak a vezikulák és a célmembrán fúziójának specifikus és hatékony irányításában.
Transzportfolyamatok szabályozása és jelentősége
A sejtek transzportfolyamatai rendkívül szigorúan szabályozottak, mivel a belső környezet állandósága, a homeosztázis fenntartása létfontosságú. A szabályozás számos szinten történhet, a génexpressziótól kezdve a fehérjék poszt-transzlációs módosításáig.
Hormonális és idegi szabályozás
Számos hormon és neurotranszmitter befolyásolja a transzporterek és csatornák aktivitását. Például az inzulin serkenti a GLUT4 glükóz transzporterek beépülését az izom- és zsírsejtek membránjába, fokozva ezzel a glükóz felvételét. Az idegrendszerben a neurotranszmitterek kötődése ligandfüggő ioncsatornákat nyit meg vagy zár be, alapvetően befolyásolva az idegimpulzusok terjedését.
A vese működésében az aldoszteron hormon befolyásolja a nátrium- és káliumtranszportot, míg az antidiuretikus hormon (ADH) az aquaporinok beépülését szabályozza a vesetubulusokba, befolyásolva a vízvisszaszívást.
Poszt-transzlációs módosítások
A transzporter fehérjék aktivitását gyakran befolyásolják poszt-transzlációs módosítások, mint például a foszforiláció. A fehérje kinázok foszfátcsoportokat adhatnak hozzá a transzporterekhez, ami megváltoztathatja konformációjukat és aktivitásukat. Ez egy gyors és hatékony módja a sejt számára, hogy reagáljon a változó körülményekre.
Például a CFTR (Cisztás Fibrózis Transzmembrán Konduktancia Regulátor) kloridcsatorna aktivitását a cAMP-függő protein kináz (PKA) foszforilációja szabályozza. Ennek a csatornának a hibás működése okozza a cisztás fibrózis betegséget.
Génexpresszió és transzporterek számának változása
Hosszabb távon a sejt szabályozhatja a transzporter fehérjék mennyiségét a membránban a génexpresszió szabályozásával. Például tartósan magas glükózszint esetén a sejtek növelhetik a glükóz transzporterek szintézisét. Ez a mechanizmus teszi lehetővé a sejtek számára, hogy alkalmazkodjanak a környezeti változásokhoz és fenntartsák a metabolikus egyensúlyt.
Transzporttal kapcsolatos betegségek és farmakológiai vonatkozások
A transzportfolyamatok zavarai számos súlyos betegséghez vezethetnek, és éppen ezért a transzporterek és ioncsatornák fontos gyógyszercélpontok. A modern orvostudomány számos terápiás megközelítése alapul ezen mechanizmusok modulálásán.
Cisztás fibrózis (CF)
A cisztás fibrózis egy örökletes betegség, amelyet a CFTR (Cisztás Fibrózis Transzmembrán Konduktancia Regulátor) gén mutációja okoz. A CFTR egy kloridion-csatorna, amely a hámsejtek membránjában található, és a kloridionok transzportjáért felelős. A hibás CFTR csatorna miatt a kloridionok nem jutnak ki megfelelően a sejtekből, ami a nyálkahártyákban (légutak, hasnyálmirigy, bél) sűrű, tapadós nyák felhalmozódásához vezet. Ez légzési problémákat, emésztési zavarokat és egyéb szövődményeket okoz.
Cukorbetegség (diabetes mellitus)
A 2-es típusú cukorbetegségben gyakran előfordul az inzulinrezisztencia, ahol az izom- és zsírsejtek nem reagálnak megfelelően az inzulinra, és a GLUT4 transzporterek nem épülnek be hatékonyan a membránba. Ennek következtében a glükóz felvétele a vérből csökken, ami magas vércukorszinthez vezet. Az SGLT2 gátlók, mint például a dapagliflozin, a vesékben gátolják az SGLT2 transzportert, csökkentve a glükóz visszaszívását és növelve a vizelettel történő kiválasztását, ezzel csökkentve a vércukorszintet.
Szívritmuszavarok (arrhythmiák)
A szívizomsejtekben található ioncsatornák (különösen a Na+, K+ és Ca2+ csatornák) hibás működése szívritmuszavarokhoz vezethet. Az úgynevezett ioncsatornopathiák genetikai mutációk miatt alakulhatnak ki, és a csatornák abnormális működését eredményezik. Számos antiarritmiás gyógyszer hatását ezeknek az ioncsatornáknak a modulációjával fejti ki, például blokkolva bizonyos nátrium- vagy káliumcsatornákat.
Magas vérnyomás (hypertonia)
A vesében található transzporterek, amelyek a nátrium és a víz reabszorpciójáért felelősek, fontos célpontjai a diuretikumoknak. Ezek a gyógyszerek gátolják a nátrium visszaszívását, ami fokozott vízürítést és csökkent vérvolument eredményez, ezáltal csökkentve a vérnyomást. Például a tiazid diuretikumok a Na+/Cl- ko-transzportert gátolják a vesetubulusokban.
Gyomorfekély és reflux
A gyomor parietális sejtjeiben található H+/K+-ATPáz pumpa, más néven proton pumpa, felelős a gyomorsav (HCl) kiválasztásáért. A proton pumpa gátlók (PPI-k), mint az omeprazol vagy pantoprazol, irreverzibilisen gátolják ezt a pumpát, drámaian csökkentve a savtermelést. Ez a gyógyszercsoport széles körben alkalmazott gyomorfekély, refluxbetegség és más gyomorsavval kapcsolatos rendellenességek kezelésére.
A transzportfolyamatok megértése nem csupán az alapvető biológiai működés kulcsa, hanem számos súlyos betegség diagnosztizálásának és terápiájának alapja is.
A transzport szerepe az idegrendszerben

Az idegrendszer, különösen az idegsejtek (neuronok) működése elképzelhetetlen a precízen szabályozott iontranszport nélkül. Az idegimpulzusok, azaz az akciós potenciálok kialakulása és terjedése teljes mértékben az ioncsatornák és pumpák összehangolt működésén alapul.
A nyugalmi állapotban lévő neuron membránja egy nyugalmi membránpotenciált tart fenn, amely a sejt belsejét negatívabbá teszi a külsejéhez képest. Ezt elsősorban a Na+/K+-ATPáz pumpa működése és a kálium-szivárgási csatornák hozzák létre, amelyek lehetővé teszik a K+ ionok kiáramlását a sejtből.
Amikor egy neuron ingert kap, a membránpotenciál megváltozik, és ha eléri a küszöbértéket, feszültségfüggő nátriumcsatornák nyílnak meg, ami Na+ ionok gyors beáramlását okozza. Ez a membrán depolarizációjához, azaz a membránpotenciál pozitív irányú eltolódásához vezet, kialakítva az akciós potenciál felszálló ágát.
Ezt követően a nátriumcsatornák inaktiválódnak, és feszültségfüggő káliumcsatornák nyílnak meg, amelyek K+ ionok kiáramlását okozzák. Ez a repolarizációhoz, majd hiperpolarizációhoz vezet, visszaállítva a nyugalmi membránpotenciált. Az akciós potenciál végigterjed az axonon, lehetővé téve az információ gyors továbbítását.
A szinapszisokban, ahol az idegsejtek kommunikálnak, a neurotranszmitterek felszabadulása exocitózissal történik. A preszinaptikus terminálba érkező akciós potenciál feszültségfüggő kalciumcsatornákat nyit meg, a Ca2+ beáramlása pedig kiváltja a neurotranszmittert tartalmazó vezikulák fúzióját a preszinaptikus membránnal és a neurotranszmitterek szinaptikus résbe ürítését. A posztszinaptikus membránon a neurotranszmitterek ligandfüggő ioncsatornákhoz kötődnek, megváltoztatva azok permeabilitását és a posztszinaptikus sejt membránpotenciálját.
Transzport a növényi sejtekben
Bár a fókusz gyakran az állati sejteken van, a növényi sejtekben is rendkívül komplex és létfontosságú transzportfolyamatok zajlanak. A növényeknek vizet, ásványi anyagokat és fotoszintetizált cukrokat kell szállítaniuk a gyökerektől a levelekig és vissza.
A növényi sejtekben a proton pumpák (H+-ATPázok) különösen fontosak. Ezek a pumpák hidrogénionokat pumpálnak ki a citoplazmából a sejtfalon kívüli térbe vagy a vakuólumba, létrehozva egy proton gradienst. Ez a gradiens hajtóereje számos szekunder aktív transzporternek, amelyek a tápanyagok, például a cukrok (szacharóz) vagy aminosavak felvételéért felelősek a gyökérsejtekbe, vagy a floémbe való betöltésükért.
Az aquaporinok szintén kulcsszerepet játszanak a növények vízháztartásában, lehetővé téve a víz gyors mozgását a sejtek között és a gyökerektől a levelekig. A növények a turgornyomás fenntartásával képesek megtartani merevségüket, ami az ozmózis és a sejtfal kölcsönhatásának eredménye. Az ionok felvétele a talajból is szigorúan szabályozott, specifikus transzporterek és csatornák segítségével.
A transzportfolyamatok kutatási módszerei
A sejtek transzportmechanizmusainak megértése hatalmas előrelépést hozott a biológia és az orvostudomány területén. Ehhez számos kifinomult kutatási módszerre volt és van szükség.
Patch-clamp technika
A patch-clamp technika egy elektrofiziológiai módszer, amely lehetővé teszi az egyes ioncsatornák áramának mérését, vagy akár egy egész sejt membránpotenciáljának és áramainak vizsgálatát. Egy rendkívül finom üvegelektródát (pipettát) illesztenek a sejtmembránhoz, és áramkörbe kapcsolják. Ez a technika forradalmasította az ioncsatornák működésének megértését, és alapvető volt az akciós potenciál részletes elemzésében.
Radioaktív izotópok és fluoreszcens jelölők
A radioaktív izotópokkal (pl. 3H-glükóz, 22Na+) vagy fluoreszcens jelölőkkel ellátott molekulák felhasználása lehetővé teszi a transzportfolyamatok nyomon követését valós időben. A sejtekbe bejutó vagy onnan kiáramló jelölt molekulák mennyisége mérhető, ami információt szolgáltat a transzport sebességéről és a transzporterek specificitásáról.
Vezikula fúziós vizsgálatok
Az exocitózis és az endocitózis vezikuláris folyamatainak tanulmányozására gyakran használnak olyan módszereket, amelyek a vezikulák és a membránok fúzióját vizsgálják. Ez magában foglalhatja a fluoreszcens festékek inkorporálását a vezikulákba, majd a fúzió során bekövetkező jelváltozás detektálását, vagy elektronmikroszkópos vizsgálatokat a vezikulák morfológiai változásainak megfigyelésére.
Molekuláris biológiai módszerek
A molekuláris biológiai technikák, mint például a génklónozás, a mutagenezis vagy a génkiütés (knockout), lehetővé teszik a kutatók számára, hogy azonosítsák a transzporter fehérjék génjeit, módosítsák azok szerkezetét és funkcióját, vagy teljesen eltávolítsák azokat, hogy tanulmányozzák a transzporter hiányának következményeit. Ez elengedhetetlen a transzporterek szerepének megértéséhez a betegségekben és a gyógyszercélpontok azonosításához.
A transzport mechanizmusok evolúciója
A membrán transzport mechanizmusok evolúciója az élet kialakulásának kezdetéig nyúlik vissza. Az első primitív sejteknek is meg kellett oldaniuk az anyagok felvételét és kiválasztását, hogy fenntarthassák belső környezetüket a külső világtól elhatárolva. Az egyszerű diffúzióból kiindulva az evolúció során egyre komplexebb és specifikusabb transzporter rendszerek alakultak ki.
Az első ioncsatornák valószínűleg viszonylag nem-szelektívek voltak, de az idő múlásával, a gén duplikációk és mutációk révén, rendkívül specifikus és szabályozott csatornák jöttek létre. A pumpák, amelyek energiát használnak a gradiens ellenében történő transzportra, lehetővé tették a sejtek számára, hogy aktívan felhalmozzanak bizonyos anyagokat, ami kulcsfontosságú volt a bonyolultabb metabolikus útvonalak és a sejtdifferenciáció kialakulásához.
A vezikuláris transzport megjelenése az eukarióta sejtekben hatalmas ugrást jelentett, lehetővé téve a nagy molekulák, sőt egész részecskék hatékony szállítását. Ez alapozta meg a sejten belüli kompartmentalizációt, a szekréciós útvonalakat és az immunrendszer működését. A transzportfolyamatok folyamatos finomodása és specializációja tette lehetővé az élővilág sokszínűségét és komplexitását, a baktériumoktól az emberi szervezetig.
Összefoglalva, a transzporttal rendelkező cella fogalma az élet alapját képező dinamikus folyamatok egész hálózatát öleli fel. A sejtmembránon keresztüli anyagcsere, legyen az passzív vagy aktív, egyszerű diffúzió vagy vezikuláris transzport, nélkülözhetetlen a sejt túléléséhez, működéséhez és a szervezet egészének homeosztázisához. A mechanizmusok mélyreható megértése kulcsfontosságú a biológia, a gyógyászat és a gyógyszerfejlesztés területén, és folyamatosan új felfedezésekkel gazdagodik.







