
Az RNS-polimerázok, avagy ribonukleinsav-polimerázok, a molekuláris biológia egyik legfontosabb és legősibb enzimcsaládját alkotják. Ezek a komplex fehérjék felelősek a genetikai információ DNS-ről RNS-re történő átírásáért, egy olyan alapvető folyamatért, amelyet transzkripciónak nevezünk. A transzkripció a génexpresszió első lépése, amely során a DNS-ben tárolt genetikai kód egy RNS molekulába íródik át. Ez az RNS molekula aztán vagy közvetlenül funkcionális szerepet tölt be a sejtben (például transzfer RNS – tRNS, riboszomális RNS – rRNS), vagy messenger RNS-ként (mRNS) szolgál a fehérjeszintézis templátjaként. Az RNS-polimerázok precíz és szabályozott működése nélkülözhetetlen az élet minden formájában, a baktériumoktól az emberig, biztosítva a sejtek megfelelő működéséhez szükséges fehérjék és RNS-molekulák előállítását.
A transzkripció folyamatának megértése kulcsfontosságú a génszabályozás, a sejtfejlődés, a betegségek kialakulása és a gyógyszerfejlesztés szempontjából. Az RNS-polimerázok a DNS kettős spiráljának egy szakaszát olvassák le, és egy komplementer RNS-szálat szintetizálnak. Ez a folyamat rendkívül komplex és szigorúan szabályozott, számos segédfehérje és szabályozó elem bevonásával történik, amelyek biztosítják, hogy a megfelelő gének a megfelelő időben és mennyiségben fejeződjenek ki. A továbbiakban részletesen megvizsgáljuk az RNS-polimerázok szerkezetét, működési mechanizmusait, a különböző típusait prokariótákban és eukariótákban, valamint a génexpresszióban betöltött kritikus szerepüket.
A transzkripció folyamata: áttekintés
A transzkripció a genetikai információ áramlásának alapvető lépése, amely a centrális dogma része. Ez a dogma kimondja, hogy a genetikai információ a DNS-ből az RNS-be, majd az RNS-ből a fehérjébe áramlik. A transzkripció során a DNS-ben tárolt genetikai információ egy RNS molekulába íródik át. Ez a folyamat alapvetően három fő szakaszra osztható: iniciációra (kezdet), elongációra (nyújtás) és terminációra (befejezés).
Az iniciáció során az RNS-polimeráz és a hozzá kapcsolódó faktorok felismerik a DNS-templát egy specifikus régióját, az úgynevezett promótert. A promóterek a gén elején helyezkednek el, és jelzik az RNS-polimeráznak, hogy hol kezdje meg az RNS szintézisét. Miután az RNS-polimeráz kötődik a promóterhez, a DNS kettős spirálja lokálisan feltekeredik, létrehozva egy transzkripciós buborékot, amely hozzáférést biztosít a DNS-szálakhoz.
Az elongáció szakaszában az RNS-polimeráz végighalad a DNS-templát szálon, miközben ribonukleotid-trifoszfátokat (NTP-ket) épít be az újonnan szintetizálódó RNS-szálba. Az RNS-polimeráz a DNS egyik szálát használja templátként (az úgynevezett templát szálat vagy antiszensz szálat), és komplementer bázisokat illeszt hozzá: adeninhez uracilt, timinhez adenint, guaninhez citozint, citozinhez guanint. Az RNS szintézise mindig 5’→3′ irányban történik.
Végül, a termináció során az RNS-polimeráz elér egy specifikus DNS-szekvenciát, a terminátort, amely jelzi a transzkripció befejezését. Ekkor az RNS-polimeráz disszociál a DNS-ről, és a frissen szintetizált RNS molekula felszabadul. A transzkripció befejezése után az RNS molekula további módosításokon eshet át, különösen eukariótákban, mielőtt betöltené funkcióját vagy felhasználódna a fehérjeszintézisben.
Az RNS-polimerázok szerkezete és működési elve
Az RNS-polimerázok rendkívül komplex, multisubunit-os enzimek, amelyek katalitikus aktivitásuk révén felelősek a ribonukleotidok polimerizációjáért. Bár a prokarióta és eukarióta RNS-polimerázok között jelentős különbségek vannak a méretben és a komplexitásban, számos alapvető szerkezeti és működési elvük közös. Mindegyik RNS-polimeráz rendelkezik egy mély hasadékkal vagy csatornával, amelyen keresztül a DNS-templát áthalad, és egy aktív hellyel, ahol a ribonukleotidok kapcsolódnak egymáshoz.
Az RNS-polimerázoknak nincs szükségük primerre az RNS szintézisének megkezdéséhez, ellentétben a DNS-polimerázokkal. Ez egy kulcsfontosságú különbség, amely lehetővé teszi számukra, hogy közvetlenül a promóter régióban kezdeményezzék a transzkripciót. Az enzim aktív centruma egy vagy több fémiont, általában magnéziumot tartalmaz, amely elengedhetetlen a katalitikus reakcióhoz, azaz a foszfodiészter kötések kialakításához a ribonukleotidok között.
Az enzim mozgása a DNS-templáton keresztül úgynevezett léptető mechanizmussal történik. Minden egyes hozzáadott nukleotid után az RNS-polimeráz egy bázispárral előrehalad a DNS-en, miközben a DNS kettős spirálja előtt feltekeredik, mögötte pedig visszatekeredik. Ez a folyamat egyidejűleg biztosítja a DNS-szálak szétválasztását az aktív helyen és az újonnan szintetizált RNS-szál leválását a templátról.
Az RNS-polimerázoknak van egy bizonyos fokú korrektúrázó képességük is, bár ez általában kevésbé hatékony, mint a DNS-polimerázok esetében. A hibásan beépített nukleotidok felismerhetők és eltávolíthatók, ami hozzájárul a transzkripció pontosságához. Ennek ellenére az RNS szintézis hibarátja magasabb, mint a DNS replikációé, ami elfogadható, mivel az RNS molekulák élettartama általában rövidebb, és sok másolat készülhet belőlük.
A prokarióta RNS-polimeráz: szerkezet és mechanizmus
A prokarióta szervezetekben, például a baktériumokban, általában egyetlen típusú RNS-polimeráz végzi az összes RNS szintézisét (mRNS, tRNS, rRNS). Ez az enzim egy rendkívül komplex, multisubunit-os fehérje, amely két fő formában létezik: mint core enzim és mint holoenzim.
A core enzim öt alegységből áll: két α (alfa) alegységből, egy β (béta) alegységből, egy β’ (béta-prim) alegységből és egy ω (omega) alegységből (α2ββ’ω). Az α alegységek a szerelési folyamatban és a promóter-kötő fehérjékkel való kölcsönhatásban játszanak szerepet. A β és β’ alegységek alkotják az enzim katalitikus magját, tartalmazzák az aktív helyet és felelősek a DNS-kötésért, valamint a nukleotidok polimerizációjáért. Az ω alegység szerepe kevésbé tisztázott, de úgy vélik, hogy az enzim stabilitásában és szerelési folyamatában van szerepe.
A holoenzim akkor keletkezik, amikor a core enzimhez egy sigma (σ) faktor kapcsolódik. A sigma faktor kulcsfontosságú az iniciációban, mivel felelős a promóter régiók specifikus felismeréséért a DNS-en. Különböző sigma faktorok léteznek, amelyek különböző géncsoportok transzkripcióját irányítják, lehetővé téve a baktériumok számára, hogy reagáljanak a környezeti változásokra vagy szabályozzák a fejlődési programokat. A sigma faktor segít az RNS-polimeráznak a helyes startpont megtalálásában, ezáltal biztosítva a génexpresszió pontosságát.
A prokarióta RNS-polimeráz holoenzimje a gének precíz felismerésének és az RNS szintézis iniciálásának mestere, a sigma faktor vezetésével.
Amikor a holoenzim kötődik a promóterhez, az úgynevezett zárt komplex képződik. Ezt követően a DNS kettős spirálja feltekeredik, egy nyitott komplexet hozva létre, amelyben a templát és a kódoló szálak szétválnak egy rövid szakaszon. Ez a transzkripciós buborék lehetővé teszi az RNS-polimeráz számára, hogy hozzáférjen a templát szálhoz és megkezdje az RNS szintézisét. Az iniciáció során gyakran előfordul az úgynevezett abortív iniciáció, amikor az RNS-polimeráz rövid, inproduktív RNS-szálakat szintetizál, mielőtt stabil elongációs komplexet alakítana ki és elindulna a promóterről.
A transzkripció fázisai prokariótákban: iniciáció, elongáció, termináció
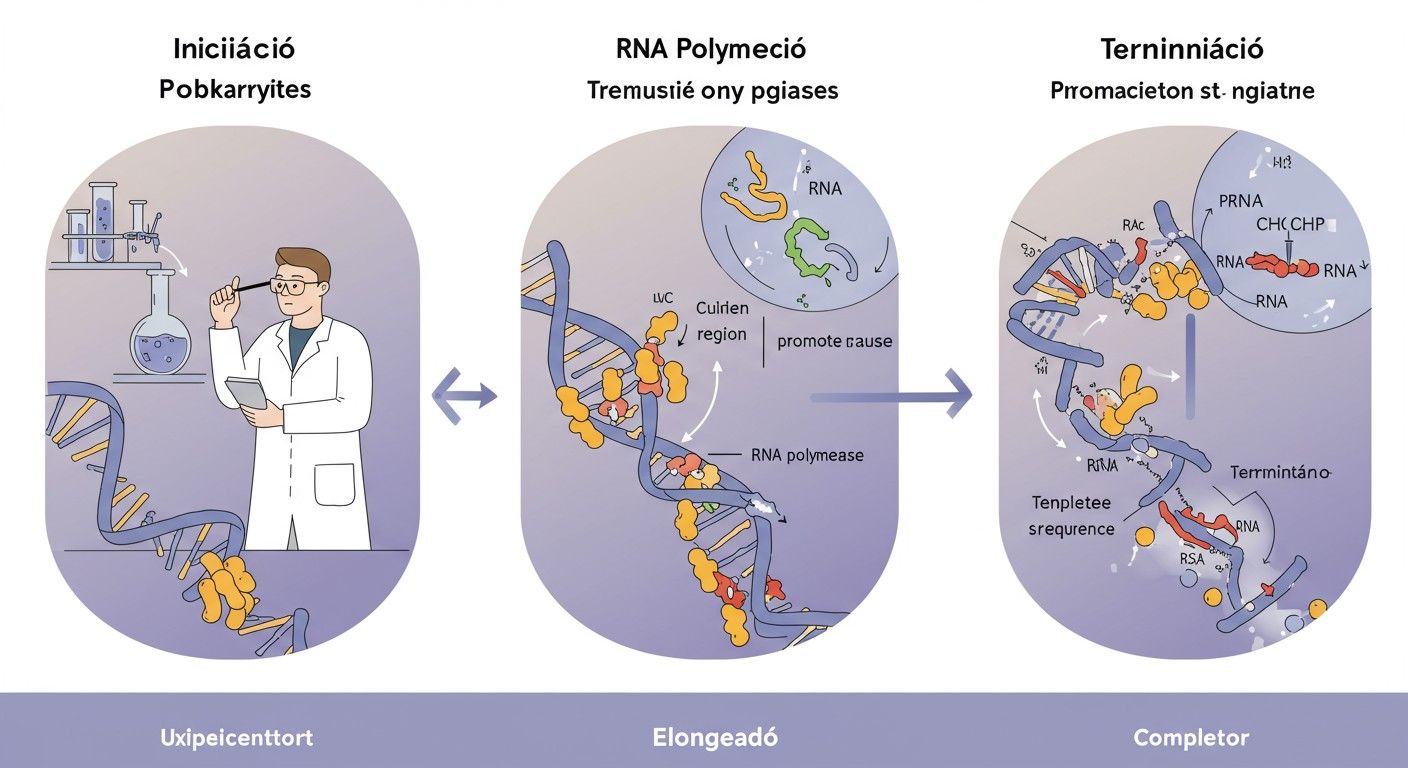
A prokarióta transzkripció három fő fázisra bontható, amelyek mindegyike szigorúan szabályozott és koordinált.
Iniciáció
Az iniciáció a transzkripció leginkább szabályozott lépése. Az RNS-polimeráz holoenzim specifikusan kötődik a promóter régióhoz, amely általában -35 és -10 bázispárral a transzkripció startpontja előtt található. Két konzervált szekvencia, a -35 régió (TTGACA) és a -10 régió (TATAAT, más néven Pribnow doboz) kulcsfontosságú a sigma faktor általi felismerésben. Ezek a szekvenciák biztosítják, hogy az RNS-polimeráz a megfelelő helyen és megfelelő irányban kezdje meg az RNS szintézisét.
A promóterhez való kötődés után az RNS-polimeráz a DNS-t feltekercseli, egy körülbelül 12-14 bázispárból álló transzkripciós buborékot hozva létre. Ez a nyitott komplex képződése energiát igényel, de nem hidrolizál ATP-t. Miután az első ribonukleotidok beépültek, és az RNS-szál hossza eléri a 8-10 nukleotidot, a sigma faktor általában disszociál a core enzimről. Ez a lépés jelzi az átmenetet az iniciációból az elongációba, és lehetővé teszi az RNS-polimeráz számára, hogy hatékonyan mozogjon a DNS-templáton.
Elongáció
Az elongáció során a core RNS-polimeráz enzim stabilan kötődik a DNS-hez, és folyamatosan halad végig a templát szálon, miközben NTP-ket épít be az újonnan szintetizálódó RNS-szálba. Az RNS szintézise 5’→3′ irányban történik, és a templát szál 3’→5′ irányban olvasódik le. Az RNS-polimeráz a DNS-t előtt feltekercseli, és mögötte visszatekercseli, fenntartva a transzkripciós buborékot. A hibásan beépített nukleotidok bizonyos mértékig korrigálhatók, bár a prokarióta RNS-polimeráz hibajavító mechanizmusa kevésbé robusztus, mint a DNS-polimerázoké.
Az elongáció sebessége változó, és számos tényező befolyásolhatja, például a DNS-szekvencia, a nukleotid-koncentráció és a kísérőfehérjék jelenléte. Bizonyos fehérjék, mint például a NusA és a NusG, kötődhetnek az elongáló RNS-polimerázhoz, és befolyásolhatják annak sebességét és folyamatosságát. Ezek a faktorok különösen fontosak az operonok transzkripciójában, ahol a génexpresszió koordinált szabályozása szükséges.
Termináció
A transzkripció terminációja akkor következik be, amikor az RNS-polimeráz elér egy specifikus DNS-szekvenciát, a terminátort. Prokariótákban két fő típusú terminációs mechanizmus létezik:
- Rho-független (intrinszik) termináció: Ez a mechanizmus a frissen szintetizált RNS-szálban lévő specifikus szekvenciákra támaszkodik. Jellemzően egy GC-gazdag, palindrom szekvenciából áll, amelyet egy uracilban gazdag régió követ. A palindrom szekvencia hajtűhurkot képez az RNS-en belül, ami destabilizálja az RNS-polimerázt. A hajtűhurok és az uracilban gazdag régió közötti gyenge rRNS-DNS hibrid kötések felszakadnak, és az RNS-polimeráz disszociál a DNS-ről.
- Rho-függő termináció: Ez a mechanizmus a Rho fehérje nevű hexamer helikázra támaszkodik. A Rho fehérje az újonnan szintetizált RNS-hez kötődik egy specifikus szekvencia, a rho felismerő hely (rut site) közelében. Az RNS-polimeráz lassulását kihasználva a Rho fehérje az RNS-en haladva utoléri az RNS-polimerázt, majd ATP hidrolízis segítségével szétválasztja az RNS-DNS hibridet, és felszabadítja az RNS-szálat.
Mindkét terminációs mechanizmus biztosítja, hogy a transzkripció a megfelelő helyen fejeződjön be, megelőzve a felesleges RNS molekulák szintézisét és hozzájárulva a génexpresszió pontos szabályozásához.
Az eukarióta RNS-polimerázok diverzitása: Pol I, Pol II, Pol III és mások
Az eukarióta sejtekben a genetikai információ sokkal komplexebb szerkezetben, a kromatin formájában van jelen, és a transzkripciót több különböző típusú RNS-polimeráz végzi, amelyek mindegyike specifikus RNS-osztályok szintéziséért felelős. Ez a specializáció lehetővé teszi az eukarióták számára, hogy finomhangolják a génexpressziót és bonyolultabb szabályozási mechanizmusokat alkalmazzanak. Az eukarióta RNS-polimerázok nagyobbak és komplexebbek, mint prokarióta társaik, és számos alegységből állnak.
Három fő típusú eukarióta RNS-polimerázt különböztetünk meg:
- RNS-polimeráz I (Pol I): Fő feladata a riboszomális RNS (rRNS) molekulák prekurzorainak szintézise, kivéve az 5S rRNS-t.
- RNS-polimeráz II (Pol II): Felelős a messenger RNS (mRNS) prekurzorok, valamint számos kis nukleáris RNS (snRNS) és mikroRNS (miRNS) szintéziséért. Ez a polimeráz a génexpresszió központi szereplője.
- RNS-polimeráz III (Pol III): Szintetizálja a transzfer RNS-eket (tRNS), az 5S rRNS-t és néhány más kis, stabil RNS molekulát.
Ezeken a fő típusokon kívül, különösen növényekben, további RNS-polimerázokat is azonosítottak, mint például az RNS-polimeráz IV (Pol IV) és az RNS-polimeráz V (Pol V), amelyek a kis interferáló RNS-ek (siRNS) szintézisében és a kromatin szerkezet szabályozásában vesznek részt.
Az eukarióta RNS-polimerázok működése sokkal több kiegészítő fehérjét igényel, mint a prokarióták esetében. Ezeket a fehérjéket transzkripciós faktoroknak nevezzük, és kulcsszerepet játszanak a promóter felismerésében, a transzkripció iniciálásában, elongációjában és terminációjában. A transzkripciós faktorok specifikusan kötődnek a DNS-hez, vagy kölcsönhatásba lépnek az RNS-polimerázzal, finomhangolva a génexpressziót és biztosítva a szövetspecifikus vagy fejlődési stádium-specifikus génműködést.
Az eukarióta RNS-polimerázok specializált munkamegosztása és a transzkripciós faktorok komplex hálózata teszi lehetővé a magasabb rendű szervezetek rendkívül finomhangolt génexpresszióját.
| RNS-polimeráz típus | Fő termék(ek) | Elhelyezkedés | Érzékenység α-amanitinre |
|---|---|---|---|
| RNS-polimeráz I (Pol I) | Nagy rRNS prekurzor (pl. 45S pre-rRNS) | Sejtmagvacska (nukleólusz) | Nem érzékeny |
| RNS-polimeráz II (Pol II) | mRNS prekurzorok, snRNS, snoRNS, miRNS | Nukleoplazma | Nagyon érzékeny |
| RNS-polimeráz III (Pol III) | tRNS, 5S rRNS, néhány snRNS | Nukleoplazma | Közepesen érzékeny |
| RNS-polimeráz IV (Pol IV) | siRNS prekurzorok (növényekben) | Nukleoplazma | Nem érzékeny |
| RNS-polimeráz V (Pol V) | siRNS prekurzorok (növényekben), kromatin módosítás | Nukleoplazma | Nem érzékeny |
Az RNS-polimeráz I: riboszomális RNS szintézis
Az RNS-polimeráz I (Pol I) feladata az eukarióta sejtben a riboszomális RNS (rRNS) molekulák többségének szintézise. Ezek az rRNS-ek a riboszómák, a fehérjeszintézis gépezetének alapvető szerkezeti és katalitikus komponensei. A Pol I kizárólag a 45S pre-rRNS-t transzkribálja, amelyből aztán poszt-transzkripciós módosítások és hasítások révén keletkezik a 18S, 5.8S és 28S rRNS. Az 5S rRNS-t az RNS-polimeráz III szintetizálja.
A Pol I a sejtmagvacskában (nukleóluszban) található, amely egy specializált nukleáris régió, ahol a riboszóma biogenezise zajlik. Az rRNS-gének több száz, tandem módon elrendezett kópiában vannak jelen a genomban, biztosítva a nagy mennyiségű rRNS termelését, amely szükséges a sejt folyamatos fehérjeszintéziséhez. A Pol I által transzkribált rRNS-géneknek saját, specifikus promótereik vannak, amelyek különböznek a Pol II és Pol III által felismert promóterektől.
A Pol I transzkripció iniciációja is transzkripciós faktorokat igényel, de ezek a faktorok specifikusan a Pol I-hez és annak promótereihez kötődnek. A legfontosabb faktorok közé tartozik az UBF (Upstream Binding Factor) és a SL1 (Selectivity Factor 1). Az UBF dimerek formájában kötődik az rRNS promóter upstream szabályozó régiójához és a core promóterhez, elősegítve a kromatin szerkezet megnyílását. Az SL1, amely egy TBP (TATA-kötő fehérje) és több TAF (TBP-asszociált faktor) alegységből álló komplex, kötődik az UBF-hez és a core promóterhez, majd toborozza a Pol I-et a startponthoz.
A Pol I által végzett rRNS szintézis rendkívül aktív folyamat, amely a sejt metabolikus állapotától függően szigorúan szabályozott. A sejt növekedési sebessége és a tápanyagellátás közvetlenül befolyásolja a Pol I aktivitását, biztosítva, hogy a riboszómák termelése összhangban legyen a sejt fehérjeszintézisi igényeivel. A Pol I disregulációja összefüggésbe hozható bizonyos betegségekkel, például a rákkal, ahol a fokozott riboszóma biogenezis hozzájárul a gyors sejtnövekedéshez.
Az RNS-polimeráz II: a messenger RNS és más kis RNS-ek
Az RNS-polimeráz II (Pol II) az eukarióta transzkripció központi szereplője, felelős a legtöbb gén, különösen a fehérjéket kódoló gének, azaz a messenger RNS (mRNS) prekurzorainak szintéziséért. Emellett számos más fontos RNS molekulát is transzkribál, mint például a kis nukleáris RNS-eket (snRNS-ek), amelyek az RNS splicingban vesznek részt, a kis nukleóluszi RNS-eket (snoRNS-ek), amelyek az rRNS módosításában játszanak szerepet, és a mikroRNS-eket (miRNS-ek), amelyek a génexpresszió poszt-transzkripciós szabályozásában fontosak.
A Pol II egy nagyméretű, 12 alegységből álló komplex, amelynek legjellemzőbb vonása a legnagyobb alegységének C-terminális doménje (CTD). A CTD egy ismétlődő heptapeptid szekvenciából áll (Tyr-Ser-Pro-Thr-Ser-Pro-Ser), amelynek foszforilációs állapota kulcsszerepet játszik a transzkripció különböző fázisainak szabályozásában és a poszt-transzkripciós módosítások, például a splicing és a poliadenilezés koordinálásában. Az α-amanitin nevű gomba toxin nagyon érzékeny a Pol II-re, ami segített a Pol II azonosításában és tanulmányozásában.
A Pol II által transzkribált gének promóterei rendkívül változatosak és komplexek. A legtöbb Pol II promóter tartalmaz egy TATA-boxot, egy AT-gazdag szekvenciát (TATA(A/T)A(A/T)), amely körülbelül -25 és -30 bázispárral a transzkripció startpontja előtt található. Azonban sok génnek nincs TATA-boxa, és más core promóter elemekre támaszkodik, mint például az iniciátor elemre (Inr) vagy a downstream promóter elemre (DPE).
Az eukarióta transzkripciós iniciáció komplexitása: általános transzkripciós faktorok

Az RNS-polimeráz II által végzett transzkripció iniciációja sokkal komplexebb, mint a prokariótáké, és számos általános transzkripciós faktor (GTF) bevonását igényli. Ezek a faktorok nem specifikusak egyetlen génre sem, hanem minden Pol II által transzkribált gén iniciációjához szükségesek. A GTF-ek szekvenciális módon kötődnek a promóterhez, és egy preiniciációs komplexet (PIC) hoznak létre, amely megfelelő platformot biztosít a Pol II számára.
A PIC felépülésének lépései a következők:
- TFIID (Transcription Factor II D): Ez a komplex az első, amely a TATA-boxhoz kötődik. A TFIID kulcsfontosságú alegysége a TBP (TATA-kötő fehérje), amely specifikusan felismeri és kötődik a TATA-boxhoz, lokális DNS-hajlítást okozva. A TBP-hez számos TAF (TBP-asszociált faktor) is kapcsolódik, amelyek a promóter más elemeinek felismerésében és a szabályozó fehérjékkel való kölcsönhatásban játszanak szerepet.
- TFIIB (Transcription Factor II B): A TFIID kötődése után a TFIIB kapcsolódik a TBP-hez és a promóter szekvenciájához. A TFIIB egy hídként funkcionál a TBP és a Pol II között, és segít meghatározni a transzkripció startpontját.
- TFIIF és RNS-polimeráz II: A TFIIF, amely a Pol II-vel előkomplexet alkot, ezt követően kötődik a PIC-hez.
- TFIIE és TFIIH: Végül a TFIIE és a TFIIH kapcsolódik a komplexhez. A TFIIH egy multisubunit-os komplex, amely helikáz és kináz aktivitással is rendelkezik. A helikáz aktivitása felelős a DNS szálak feltekercseléséért a transzkripciós buborék kialakításához, míg a kináz aktivitása foszforilálja a Pol II CTD-jét, ami elengedhetetlen az iniciációról az elongációra való áttéréshez.
Miután a PIC teljesen összeállt, és a Pol II CTD-je foszforilálódott, az RNS-polimeráz elindul a promóterről, és megkezdődik az elongáció. Az iniciáció szabályozása rendkívül összetett, és nem csak a GTF-ekre, hanem számos génspecifikus transzkripciós aktivátorra és represszorra, valamint a kromatin szerkezetre is támaszkodik.
Az RNS-polimeráz II elongációja és a kísérőfehérjék szerepe
Az RNS-polimeráz II által végzett elongáció egy dinamikus és szigorúan szabályozott folyamat, amely során az enzim végighalad a DNS-templáton, és az RNS-szálat szintetizálja. Az elongáció során az RNS-polimeráz II-höz számos elongációs faktor kötődik, amelyek stabilizálják az elongációs komplexet, növelik az enzim folyamatosságát, és segítik a nukleoszómákon való áthaladást. A nukleoszómák a DNS-t tartalmazó hiszton fehérjékből álló szerkezeti egységek, amelyek akadályt jelenthetnek a transzkripció számára.
A Pol II CTD-jének foszforilációs mintázata kritikus szerepet játszik az elongáció szabályozásában. Különböző szerin reziduók foszforilációja jelzi a transzkripció különböző fázisait és toborozza a megfelelő feldolgozó enzimeket. Például a Ser5 foszforilációja az iniciációhoz és a 5′ sapka (cap) hozzáadásához kapcsolódik, míg a Ser2 foszforilációja az elongációhoz és a 3′ vég hasításához, valamint a poliadenilezéshez.
Az elongációs faktorok közé tartozik például a NELF (Negative Elongation Factor) és a DSIF (DRB Sensitivity-Inducing Factor), amelyek kezdetben gátolhatják az elongációt, lehetővé téve a Pol II számára, hogy rövid RNS-szálakat szintetizáljon, mielőtt teljes hosszúságú transzkriptumot hozna létre. Ez a „promóter-proximalis leállás” egy fontos szabályozási pont, amely lehetővé teszi a génexpresszió gyors bekapcsolását a megfelelő jel hatására.
A P-TEFb (Positive Transcription Elongation Factor b) kináz komplex foszforilálja a NELF-et és a DSIF-et, valamint a Pol II CTD-jének Ser2-jét, ezzel feloldva az elongációs gátat és lehetővé téve a Pol II számára, hogy folytassa a transzkripciót. Ez a mechanizmus kulcsfontosságú a gyorsan reagáló gének szabályozásában, például a hőstressz-válasz vagy az immunválasz során.
Az elongáció során a kromatin szerkezet is dinamikusan változik. A kromatin remodelling komplexek és a hiszton módosító enzimek együttműködnek az RNS-polimerázzal, hogy megnyissák a kromatin szerkezetet a transzkripciós buborék előtt, és visszaállítsák azt az enzim áthaladása után. Ez a folyamat biztosítja, hogy a DNS hozzáférhető legyen a Pol II számára, miközben fenntartja a genom integritását és a kromatin megfelelő tömörítését.
Az RNS-polimeráz II terminációja és a poszt-transzkripciós módosítások
Az RNS-polimeráz II által végzett transzkripció terminációja szorosan összekapcsolódik az újonnan szintetizált mRNS prekurzorok poszt-transzkripciós módosításaival. Eukariótákban az mRNS molekulák számos átalakításon esnek át, mielőtt elhagynák a sejtmagot és részt vennének a fehérjeszintézisben. Ezek a módosítások magukban foglalják az 5′ sapka (cap) hozzáadását, az intronok eltávolítását splicing révén, és a 3′ poli(A) farok hozzáadását.
A Pol II terminációjának két fő modellje létezik:
- Alloszterikus modell: Ez a modell feltételezi, hogy az RNS-polimeráz II egy specifikus terminációs szekvencia elérésekor konformációs változásokon megy keresztül, ami csökkenti a DNS-hez való affinitását és destabilizálja az elongációs komplexet.
- Torpedó modell: Ez a modell szélesebb körben elfogadott. A Pol II a transzkripció során egy poliadenilezési szignál (poly(A) signal) szekvenciát transzkribál az mRNS-be. Ennek a szignálnak a felismerése számos enzim toborzásához vezet, amelyek hasítják az RNS-szálat a poli(A) szignál után, és hozzáadják a poli(A) farkat. Azonban az RNS-polimeráz II gyakran tovább transzkribálja a hasítási pont után. Ebben a modellben egy exonukleáz (pl. Xrn2) kötődik a hasított, nem poliadenilezett RNS-véghez, és „torpedóként” halad az elongáló Pol II felé, lebontva az RNS-t. Amikor az exonukleáz eléri az RNS-polimerázt, destabilizálja azt, és a Pol II disszociál a DNS-ről.
A 5′ sapka (egy 7-metilguanozin nukleotid, amely fordított orientációban kapcsolódik az RNS 5′ végéhez) korán, az elongáció kezdetén hozzáadódik. Ez a sapka védi az mRNS-t a lebomlástól, és fontos szerepet játszik a riboszóma kötődésében és a transzláció iniciációjában. A sapka hozzáadásáért felelős enzimek a Pol II CTD-jének foszforilációjától függően toborzódnak.
A splicing, azaz az intronok eltávolítása és az exonok összeillesztése, gyakran ko-transzkripciós folyamat, ami azt jelenti, hogy még azelőtt megtörténik, hogy a transzkripció befejeződne. A spliceoszóma, egy komplex ribonukleoprotein gépezet, felismeri az intron-exon határokat, és precízen kivágja az intronokat. A splicing mechanizmusa rendkívül fontos a génexpresszió diverzitásában, mivel az alternatív splicing lehetővé teszi, hogy egyetlen génből többféle fehérje is létrejöjjön.
A poli(A) farok, egy hosszú adenin nukleotidokból álló lánc (általában 50-250 nukleotid), a 3′ vég hasítása után adódik hozzá az mRNS-hez. Ez a farok védi az mRNS-t a lebomlástól, segíti a sejtmagból való exportját, és befolyásolja a transzláció hatékonyságát. A poli(A) farok hozzáadásáért felelős enzimek is a Pol II CTD-jéhez kötődnek, koordinálva a transzkripció befejezését és az mRNS feldolgozását.
Az RNS-polimeráz III: transzfer RNS, 5S rRNS és egyéb kis RNS-ek
Az RNS-polimeráz III (Pol III) egy másik kulcsfontosságú eukarióta RNS-polimeráz, amely a transzfer RNS-ek (tRNS), az 5S riboszomális RNS (5S rRNS) és számos más kis, stabil RNS molekula (pl. U6 snRNS, 7SL RNS) szintéziséért felelős. Ezek az RNS-ek létfontosságú szerepet játszanak a fehérjeszintézisben, a splicingban és a sejt más alapvető folyamataiban.
A tRNS-ek felelősek az aminosavak szállításáért a riboszómákhoz a transzláció során, míg az 5S rRNS a riboszóma egyik szerkezeti komponense. A Pol III által transzkribált RNS-ek általában rövid, jól definiált molekulák, amelyek nem igényelnek olyan kiterjedt poszt-transzkripciós feldolgozást, mint az mRNS-ek, bár a tRNS-ek érése is magában foglalja az intronok eltávolítását és specifikus bázismódosításokat.
A Pol III promóterei egyedülállóak, mivel gyakran a kódoló régió belül találhatóak, nem pedig upstream. Ez azt jelenti, hogy a transzkripciós faktorok a génen belül kötődnek, és nem a startpont előtt. Például a tRNS-gének promóterei két konzervált boxot, az A és B boxot tartalmazzák, míg az 5S rRNS-gének promótere egy C boxot tartalmaz. Ezeket a belső promótereket a Pol III-specifikus transzkripciós faktorok, mint például a TFIIIA, TFIIIB és TFIIIC, ismerik fel.
- TFIIIA: Az 5S rRNS gén C boxához kötődik, és segít a TFIIIC toborzásában.
- TFIIIC: Kötődik a tRNS gének A és B boxaihoz, valamint a TFIIIA-hoz (5S rRNS gén esetén).
- TFIIIB: Ez a faktor a TBP-t is tartalmazza, de specifikusan a Pol III promótereihez kötődik a TFIIIC (vagy TFIIIA és TFIIIC) segítségével, és toborozza a Pol III-at.
A Pol III iniciációja is egy preiniciációs komplex felépítését foglalja magában, amely a Pol III-at a megfelelő startponthoz vezeti. A Pol III terminációja is egyszerűbb, mint a Pol II esetében, és gyakran egy rövid, T-gazdag szekvencia elérésével történik, amely destabilizálja az elongációs komplexet.
A Pol III aktivitása, hasonlóan a Pol I-hez, szigorúan szabályozott, és a sejt növekedési és proliferációs állapotától függ. A megnövekedett Pol III aktivitás összefüggésbe hozható a rákos sejtek gyors növekedésével, ami potenciális terápiás célponttá teszi.
További eukarióta RNS-polimerázok: Pol IV és Pol V növényekben

A növényekben a három fő eukarióta RNS-polimerázon (Pol I, Pol II, Pol III) kívül két további, specializált RNS-polimerázt is azonosítottak: az RNS-polimeráz IV (Pol IV) és az RNS-polimeráz V (Pol V). Ezek a polimerázok kulcsszerepet játszanak a génexpresszió epigenetikai szabályozásában, különösen a RNS-függő DNS-metiláció (RdDM) útvonalában, amely a transzpozonok és a repetitív szekvenciák elhallgattatásáért felelős.
A Pol IV elsősorban a kis interferáló RNS-ek (siRNS) prekurzorait szintetizálja. Ezek a siRNS-ek aztán feldolgozódnak, és a RNS-indukált csendesítő komplex (RISC) részeként célzottan lebontják a komplementer mRNS-eket, vagy irányítják a DNS-metilációt a genomban. A Pol IV aktivitása elengedhetetlen a genom stabilitásának fenntartásához, mivel segít elhallgattatni a potenciálisan káros mobilis elemeket.
A Pol V szintén a génexpresszió elhallgattatásában játszik szerepet, de a nem kódoló RNS-ek szintéziséért felelős, amelyek a DNS-metilációt és a hiszton módosításokat irányítják a célzott kromatin régiókban. A Pol V által szintetizált RNS-ek gyakran nem stabilak, és csak közvetítőként szolgálnak a kromatin remodelling és a DNS-metiláz enzimek toborzásában. Ez a mechanizmus biztosítja a stabil és örökölhető génelhallgattatást a növényekben.
Mind a Pol IV, mind a Pol V szerkezetileg hasonlít a Pol II-re, és valószínűleg egy közös ősből fejlődtek ki. Azonban alegység-összetételükben és funkciójukban specializálódtak, hogy betöltsék egyedi szerepüket a növényi epigenetikai szabályozásban. Ezeknek a polimerázoknak a felfedezése rávilágított a génszabályozás komplexitására és a nem kódoló RNS-ek sokoldalú szerepére az eukarióta szervezetekben.
A génexpresszió szabályozása az RNS-polimerázok szintjén
A génexpresszió szabályozása rendkívül összetett és többszintű folyamat, amely biztosítja, hogy a megfelelő gének a megfelelő időben és mennyiségben fejeződjenek ki. Az RNS-polimerázok aktivitásának szabályozása a génexpresszió egyik legfontosabb ellenőrző pontja, mind prokariótákban, mind eukariótákban.
Prokariótákban a génszabályozás gyakran az operon modell alapján történik. Egy operon egy promóterből, operátor régióból és több, funkcionálisan kapcsolódó génből áll. Az operátor régióhoz kötődő represszor fehérjék gátolhatják az RNS-polimeráz kötődését vagy mozgását, ezáltal elnyomva a gének transzkripcióját. Ezzel szemben aktivátor fehérjék segíthetik az RNS-polimeráz kötődését a promóterhez, növelve a transzkripciót. A sigma faktorok diverzitása is hozzájárul a prokarióta génszabályozáshoz, lehetővé téve a különböző környezeti feltételekre való válaszokat.
A génexpresszió szabályozása az RNS-polimerázok szintjén a sejt alkalmazkodóképességének és differenciálódásának alapja, lehetővé téve a precíz válaszokat a belső és külső jelekre.
Eukariótákban a szabályozás sokkal bonyolultabb. Az RNS-polimeráz II aktivitását számos tényező befolyásolja:
- Core promóterek: A TATA-box, Inr, DPE és más elemek befolyásolják a Pol II kötődését és a transzkripció startpontját.
- Enhancerek és silencerek: Ezek a DNS-szekvenciák távol is lehetnek a gén promóterétől, de specifikus transzkripciós faktorok (aktivátorok és represszorok) kötődnek hozzájuk. Az aktivátorok kölcsönhatásba lépnek a mediátor komplexszel, amely egy nagy fehérjekomplex, ami hídként funkcionál az aktivátorok és a Pol II/GTF-ek között, serkentve a transzkripciót. A represszorok pedig gátolják azt.
- Kromatin szerkezet: A DNS eukariótákban hisztonok köré tekeredve, kromatin formájában van jelen. A kromatin tömörsége nagyban befolyásolja a gének hozzáférhetőségét az RNS-polimerázok számára. A hiszton módosítások (acetiláció, metiláció, foszforiláció) és a DNS-metiláció megváltoztathatja a kromatin szerkezetét, befolyásolva a génexpressziót.
- Elongációs kontroll: Amint azt korábban említettük, a Pol II gyakran megáll a transzkripció elején. Az elongációs faktorok és a Pol II CTD foszforilációja szabályozza ezt a megállást és a gének teljes hossza transzkripciójának feloldását.
Az RNS-polimerázok aktivitásának szabályozása tehát nem csak a promóterhez való kötődés szintjén történik, hanem az iniciáció, elongáció és termináció minden fázisában, számos kiegészítő fehérje és a kromatin szerkezet dinamikus változásainak bevonásával. Ez a komplex szabályozási hálózat teszi lehetővé a sejtek differenciálódását, a szövetek specializációját és a szervezet alkalmazkodását a változó környezeti feltételekhez.
Epigenetikai hatások és a kromatin szerkezet szerepe
Az eukarióta génexpresszió szabályozásában az epigenetikai mechanizmusok és a kromatin szerkezet rendkívül fontos szerepet játszanak. Az epigenetika olyan örökölhető változásokat jelent a génexpresszióban, amelyek nem járnak a DNS-szekvencia megváltozásával. Ezek a változások közvetlenül befolyásolják az RNS-polimerázok hozzáférését a DNS-templáthoz.
A DNS a sejtmagban hiszton fehérjék köré tekeredve, kromatin formájában van jelen. A kromatin lehet lazább, ún. eukromatin (transzkripcionálisan aktív), vagy tömöttebb, ún. heterokromatin (transzkripcionálisan inaktív). Az RNS-polimerázok csak az eukromatin régiókhoz férnek hozzá hatékonyan. A kromatin szerkezetét számos módosítás befolyásolja:
- Hiszton módosítások: A hiszton fehérjék N-terminális farkai számos kovalens módosításon eshetnek át, mint például acetiláció, metiláció, foszforiláció, ubikvitináció. Ezek a módosítások megváltoztatják a hisztonok és a DNS közötti kölcsönhatásokat, valamint affinitási helyeket hoznak létre más fehérjék számára. Például a hiszton acetiláció (általában a hiszton acetiltranszferázok, HAT-ok által végzett) lazítja a kromatin szerkezetét, elősegítve a transzkripciót, míg a hiszton deacetiláció (hiszton deacetilázok, HDAC-ok által végzett) tömöríti a kromatint és elnyomja a génexpressziót.
- DNS-metiláció: A DNS-metiláció a citozin bázisok metilációját jelenti, általában CpG dinukleotidokon. A metilált CpG szigetek a promóter régiókban gyakran a génelhallgatással járnak együtt, mivel gátolják a transzkripciós faktorok kötődését, vagy toboroznak metil-kötő fehérjéket, amelyek tömörítik a kromatint.
- Kromatin remodelling komplexek: Ezek az ATP-függő komplexek képesek átrendezni a nukleoszómákat, elmozdítva vagy eltávolítva őket, így szabaddá téve a DNS-t az RNS-polimerázok és a transzkripciós faktorok számára.
Az RNS-polimerázok és a transzkripciós gépezet szorosan együttműködik ezekkel az epigenetikai mechanizmusokkal. A transzkripciós faktorok nemcsak közvetlenül befolyásolják az RNS-polimeráz aktivitását, hanem toborozhatnak hiszton módosító enzimeket és kromatin remodelling komplexeket is a célgének promótereihez, ezzel megváltoztatva a lokális kromatin szerkezetet és befolyásolva a transzkripciót. Ez a dinamikus kölcsönhatás biztosítja a génexpresszió finomhangolását és a sejtek hosszú távú memória mechanizmusait, amelyek alapvetőek a fejlődésben és a betegségekben egyaránt.
Az RNS-polimerázok szerepe a betegségekben és a gyógyszerfejlesztésben
Az RNS-polimerázok alapvető szerepe a génexpresszióban azt jelenti, hogy működésük zavarai számos betegség kialakulásához hozzájárulhatnak. Ezen enzimek megcélzása ezért ígéretes stratégia a gyógyszerfejlesztésben.
Bakteriális RNS-polimerázok és antibiotikumok:
A bakteriális RNS-polimerázok szerkezeti és működésbeli különbségei az eukarióta polimerázokhoz képest lehetőséget adnak a szelektív gyógyszerfejlesztésre. A rifampicin egy széles spektrumú antibiotikum, amely specifikusan kötődik a bakteriális RNS-polimeráz β alegységéhez, gátolva az RNS szintézis iniciációját. Ez a gyógyszer kulcsfontosságú a tuberkulózis (TBC) kezelésében, de a rezisztencia kialakulása miatt újabb hatóanyagokra van szükség. A bakteriális RNS-polimerázok további gátlása továbbra is aktív kutatási terület.
Eukarióta RNS-polimerázok és betegségek:
Az eukarióta RNS-polimerázok diszfunkciója számos humán betegséggel hozható összefüggésbe:
- Rák: A rákos sejtek gyakran fokozottan proliferálnak, ami megnövekedett fehérjeszintézis igényt jelent. Ennek kielégítéséhez a Pol I és Pol III aktivitása gyakran túlaktivált a rákos sejtekben, ami fokozott rRNS és tRNS termeléshez vezet. Az ezeket a polimerázokat célzó gátlószerek (pl. CX-5461, amely a Pol I-et gátolja) potenciális rákellenes terápiás lehetőségeket kínálnak. A Pol II szabályozási zavarai is hozzájárulnak a tumorfejlődéshez, mivel a sejtciklust szabályozó vagy apoptózist indukáló gének expressziója megváltozhat.
- Vírusfertőzések: Sok vírus, különösen az RNS vírusok, saját RNS-polimerázokat kódolnak a replikációjukhoz. Ezeket a virális polimerázokat célzó gyógyszerek (pl. remdesivir az RNS-függő RNS-polimerázt gátolja, amely számos RNS vírus replikációjában szerepet játszik) hatékony antivirális szerekként működhetnek. Más vírusok (pl. herpeszvírusok) a gazdasejt Pol II-jét használják a génjeik transzkripciójához, kihasználva a sejt gépezetét.
- Genetikai rendellenességek: Az RNS-polimeráz alegységeket vagy a velük kölcsönható transzkripciós faktorokat kódoló gének mutációi súlyos fejlődési rendellenességekhez vagy betegségekhez vezethetnek. Például a Pol III alegységek mutációi ritka genetikai szindrómákkal (pl. leukodisztrófia) hozhatók összefüggésbe.
- Autoimmun betegségek: Bizonyos autoimmun betegségekben (pl. szisztémás szklerózis) a betegek RNS-polimeráz I és III ellen termelnek autoantitesteket, ami az enzimek működésének zavarát és a betegség patogenezisét okozhatja.
A gyógyszerfejlesztésben az RNS-polimerázok mint célpontok vizsgálata magában foglalja a szerkezetük részletes feltérképezését (röntgenkrisztallográfia, krio-elektronmikroszkópia), a gátlószerek azonosítását és optimalizálását, valamint a rezisztencia mechanizmusok megértését. A specifikus gátlószerek kifejlesztése, amelyek csak a betegséget okozó polimerázt (pl. bakteriális vagy virális) célozzák, miközben kímélik a gazdasejt polimerázait, kulcsfontosságú a mellékhatások minimalizálásához.
A transzkripciós gépezet evolúciója

Az RNS-polimerázok és a transzkripciós gépezet evolúciója az élet történetének egyik legősibb és legfontosabb folyamatát tükrözi. A DNS-ről RNS-re történő átírás képessége már az első sejtekben is jelen volt, és az evolúció során különböző szervezetekben specializálódott és diverzifikálódott.
Úgy gondolják, hogy az összes ma létező RNS-polimeráz egy közös ősi enzimtől származik. A prokarióta RNS-polimeráz (egy típus) és az eukarióta RNS-polimerázok (Pol I, Pol II, Pol III) közötti szerkezeti hasonlóságok alátámasztják ezt az elméletet. Különösen a katalitikus magot alkotó β és β’ alegységek (prokariótákban) és a legnagyobb két alegység (eukariótákban) mutat jelentős szekvenciahomológiát és szerkezeti hasonlóságot, ami arra utal, hogy ezek az alegységek az enzim ősi, alapvető működéséért felelősek.
Az evolúció során a transzkripciós gépezet komplexitása megnőtt. A prokariótákban egyetlen RNS-polimeráz elegendő az összes RNS típus szintéziséhez, míg az eukariótákban három (vagy több) specializált polimeráz alakult ki. Ez a diverzifikáció valószínűleg a genom méretének és komplexitásának növekedésével, valamint a génexpresszió finomabb szabályozásának igényével járt együtt. Például a Pol II, amely a fehérjéket kódoló gének transzkripciójáért felelős, számos további alegységet és transzkripciós faktort szerzett, amelyek lehetővé teszik a génexpresszió rendkívül precíz és kontextus-függő szabályozását.
A szigma faktorok megjelenése a prokariótákban, és az általános transzkripciós faktorok (GTF-ek) és génspecifikus transzkripciós faktorok (aktivátorok, represszorok) komplex hálózatának kialakulása az eukariótákban, mind a génszabályozás evolúciójának fontos lépései voltak. Ezek a faktorok lehetővé teszik a szervezetek számára, hogy reagáljanak a környezeti változásokra, differenciálódjanak és komplex fejlődési programokat hajtsanak végre.
A növényekben talált Pol IV és Pol V létezése tovább hangsúlyozza az RNS-polimerázok evolúciós plaszticitását és képességét, hogy új funkciókat szerezzenek a génszabályozásban, különösen az epigenetikai mechanizmusok területén. A transzkripciós gépezet folyamatos tanulmányozása segít megérteni az élet alapvető folyamatainak eredetét és evolúcióját.
A modern kutatások és a jövőbeli perspektívák
Az RNS-polimerázok kutatása a molekuláris biológia egyik legdinamikusabban fejlődő területe. A modern technológiák, mint például a magas felbontású struktúraanalízis (röntgenkrisztallográfia, krio-elektronmikroszkópia), lehetővé tették az RNS-polimerázok és a hozzájuk kapcsolódó faktorok atomi szintű szerkezetének felderítését. Ezek a szerkezeti adatok kritikusak az enzim működési mechanizmusainak, a DNS-sel és az RNS-sel való kölcsönhatásainak, valamint a transzkripciós faktorokkal való kapcsolatainak megértéséhez.
A single-molecule technikák, mint például az optikai csipeszek vagy a mágneses csipeszek, lehetővé teszik az RNS-polimeráz egyes molekuláinak viselkedésének valós idejű megfigyelését. Ezek a módszerek betekintést nyújtanak az enzim mozgásába a DNS-en, a transzkripció sebességébe, a hibajavítási mechanizmusokba és a leállások dinamikájába, amelyek a hagyományos biokémiai módszerekkel nehezen vizsgálhatók.
A genomikai és transzkriptomikai megközelítések (pl. ChIP-seq, RNA-seq, GRO-seq) forradalmasították a génexpresszió és az RNS-polimeráz eloszlásának tanulmányozását a teljes genomon. Ezek a technikák lehetővé teszik a kutatók számára, hogy azonosítsák az RNS-polimeráz kötődési helyeit, a transzkripciós startpontokat, az elongációs mintázatokat és a génszabályozásban részt vevő cisz- és transz-elemeket széles körben. A modern bioinformatikai eszközökkel kombinálva ezek az adatok segítenek feltárni a génexpressziós hálózatok komplexitását.
A CRISPR-alapú génszerkesztési technológiák, különösen a dCas9 (dezaktivált Cas9) alkalmazása, új lehetőségeket nyitnak meg az RNS-polimerázok aktivitásának célzott manipulálására. A dCas9-et fúziós fehérjékkel (pl. aktivátorokkal vagy represszorokkal) kombinálva specifikus gének transzkripciója be- vagy kikapcsolható anélkül, hogy a DNS-szekvenciát megváltoztatnánk. Ez a technológia ígéretes eszköz a génfunkciók tanulmányozásában és potenciálisan a génterápiában is.
A jövőbeli kutatások valószínűleg továbbra is a transzkripciós gépezet alapvető mechanizmusainak, a szabályozás finomhangolásának és az epigenetikai tényezőkkel való kölcsönhatásoknak a mélyebb megértésére fókuszálnak. Az RNS-polimerázok szerepének feltárása a betegségekben, különösen a rákban és a fertőző betegségekben, továbbra is kulcsfontosságú lesz a célzott terápiák és új gyógyszerek kifejlesztésében. A mesterséges intelligencia és a gépi tanulás egyre nagyobb szerepet kap a hatalmas genomikai adatok elemzésében és a komplex szabályozási mintázatok azonosításában, ami új betekintést nyújthat az RNS-polimerázok működésébe és a génexpresszió szabályozásába.







