
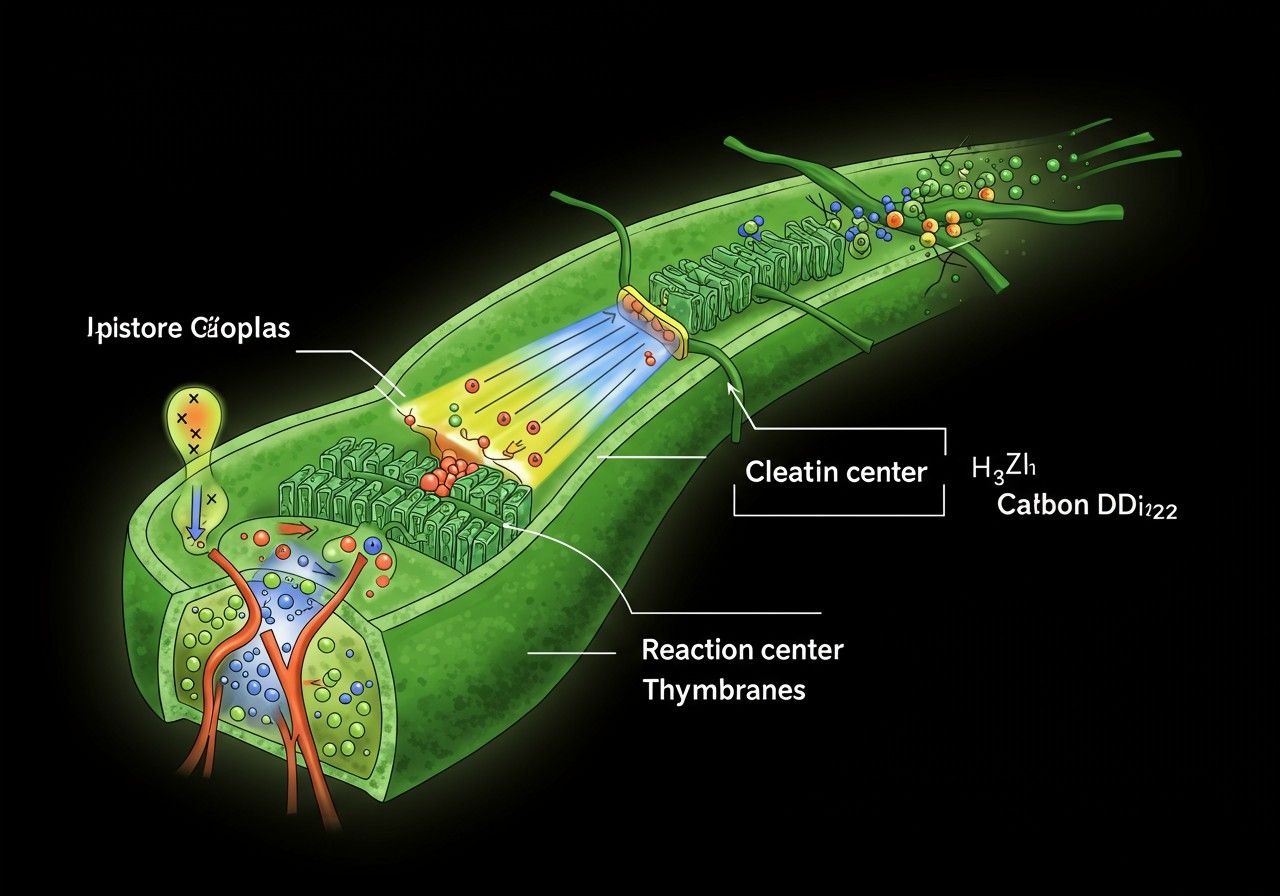
A földi élet alapját képező folyamatok közül kiemelkedik a fotoszintézis, az a lenyűgöző biokémiai mechanizmus, amely a napfény energiáját kémiai energiává alakítja. Ennek a komplex rendszernek a szívében, a fényenergia hasznosításának központi helyén található a reakciócentrum. Ez a molekuláris gépezet felelős azért a kritikus lépésért, amely elindítja az elektronok áramlását, és végül lehetővé teszi a cukrok szintézisét a szén-dioxidból. A reakciócentrum megértése kulcsfontosságú ahhoz, hogy felfogjuk a növények, algák és bizonyos baktériumok hihetetlen képességét a fényenergia átalakítására, és ezzel bolygónk ökoszisztémájának fenntartására.
A fotoszintézis nem csupán egy biológiai jelenség; ez egy olyan folyamat, amely az életet formálta a Földön, oxigénnel gazdagítva a légkört és táplálékot biztosítva a legtöbb élőlény számára. A reakciócentrum, mint a fényenergia-átalakítás elsődleges helyszíne, a legapróbb részletekig optimalizálódott az evolúció során, hogy a lehető legnagyobb hatékonysággal végezze el feladatát. Ez a cikk a reakciócentrum szerkezetébe, működésébe, típusai közé és a fotoszintézis egészében betöltött pótolhatatlan szerepébe nyújt mélyebb betekintést, feltárva ezen molekuláris csoda bonyolultságát és eleganciáját. A reakciócentrumok kutatása folyamatosan újabb és újabb felfedezéseket hoz, amelyek nemcsak az alapvető biológiai mechanizmusok megértéséhez járulnak hozzá, hanem a megújuló energiaforrások és a biotechnológia fejlesztéséhez is inspirációt adnak.
A fotoszintézis tágabb kontextusa és a fénygyűjtő komplexek
A reakciócentrumok működésének teljes megértéséhez elengedhetetlen a fotoszintézis tágabb kereteinek áttekintése. A fotoszintézis két fő szakaszra osztható: a fényfüggő reakciókra, amelyek a tilakoid membránokban zajlanak, és a fényfüggetlen reakciókra (Calvin-ciklus), amelyek a kloroplasztisz sztromájában mennek végbe. A reakciócentrumok a fényfüggő reakciók kulcsfontosságú elemei, ahol a fényenergia kémiai energiává alakul.
A fénygyűjtő komplexek (LHC – Light-Harvesting Complex) feladata a napfény fotonjainak befogása és energiájuk átadása a reakciócentrumoknak. Ezek a komplexek klorofill és karotinoid pigmentek széles skáláját tartalmazzák, amelyek különböző hullámhosszúságú fényt képesek elnyelni. Az elnyelt fény energiája rezonancia transzfer útján, rendkívül gyorsan és hatékonyan vándorol a pigmentmolekulák között, amíg el nem éri a reakciócentrumot, amely a rendszer fókuszpontjaként működik.
Ez az energiaátadás rendkívül precíz és irányított, minimalizálva az energiaveszteséget, ami a fotoszintézis rendkívüli hatékonyságát biztosítja. Képzeljünk el egy antennarendszert, amely aprólékosan hangolva van a fény befogására és továbbítására. A fénygyűjtő komplexek pontosan így működnek, koncentrálva a szétszórt fényenergiát egyetlen, kulcsfontosságú pontra: a reakciócentrumra. Ennek a hatékony energiaátadásnak köszönhetően a növények képesek maximálisan kihasználni a rendelkezésre álló fényenergiát, még alacsonyabb fényviszonyok között is, illetve alkalmazkodni a változó fényintenzitáshoz.
A fénygyűjtő komplexek nem csupán passzív antennák; dinamikusan szabályozhatók, hogy megakadályozzák a túlzott fényenergia okozta károsodást. Ez a szabályozás magában foglalja a pigmentek orientációjának és a komplexek közötti energiaátadás hatékonyságának finomhangolását, biztosítva a reakciócentrumok optimális működését a környezeti fényviszonyok széles skáláján.
A reakciócentrum definíciója és alapvető funkciója
A reakciócentrum egy speciális fehérje-pigment komplex, amely a fotoszintézis membránjaiban, például a tilakoid membránokban (növények, algák) vagy a plazmamembránban (fotoszintetikus baktériumok) található. Fő feladata a fényenergia befogása és az azt követő töltésszétválasztás, ami egy elektron transzfer folyamatot indít el. Ez a töltésszétválasztás az, ami a fényenergiát stabil kémiai energiává alakítja, elindítva a fotoszintetikus elektrontranszport láncot.
A reakciócentrum nem önmagában egyetlen molekula, hanem számos, precízen elrendezett komponensből álló makromolekuláris gépezet. Ezek közé tartoznak a speciális klorofill (vagy bakterio-klorofill) molekulák, a feofitinek (klorofill-szerű molekulák magnézium nélkül), a kinonok, és vas-kén klaszterek. Ezek a komponensek egy szigorúan meghatározott sorrendben helyezkednek el a fehérje mátrixban, biztosítva az elektronok egyirányú, rendkívül gyors és hatékony áramlását, ami elengedhetetlen a fényenergia stabil kémiai formába történő átalakításához.
A reakciócentrum legfontosabb eleme a primer elektron donor, amely általában egy speciális klorofill dimer (pl. P680 a PS II-ben, P700 a PS I-ben, vagy P870 a purpur baktériumokban). Ez a molekulapár az, amely a fénygyűjtő komplexekből érkező energiát befogadja, és gerjesztett állapotba kerülve leadja első elektronját egy akceptor molekulának. Ez a folyamat a fotoszintézis „szíve”, a pont, ahol a fényenergia valójában kémiai munkává alakul át, elindítva a komplex biokémiai kaszkádot, amely végül a szerves anyagok szintéziséhez vezet.
A reakciócentrum szerkezeti felépítése és komponensei
A reakciócentrumok szerkezete rendkívül komplex és finoman hangolt, ami lehetővé teszi számukra a rendkívül hatékony energiaátalakítást. Bár a különböző organizmusokban található reakciócentrumok között vannak eltérések, alapvető strukturális és funkcionális elvük hasonló. Általában egy transzmembrán fehérje komplexből állnak, amely számos kofaktort köt meg, és ezek a kofaktorok a fehérje vázon belül pontosan meghatározott térbeli elrendezésben helyezkednek el.
Fehérje alegységek és szerepük
A reakciócentrum vázát transzmembrán fehérjék alkotják, amelyek többszörösen átszelik a membránt. Ezek a fehérjék nem csupán egy támasztó szerkezetet biztosítanak, hanem aktívan részt vesznek a pigmentek és elektronakceptorok pozicionálásában, biztosítva az optimális távolságot és orientációt az elektronátvitelhez. Például a purpur baktériumok reakciócentrumában három fő alegység található: L, M és H. A növények fotoszisztémáiban (PS I és PS II) még több alegység alkotja a komplexet, amelyek közül néhány közvetlenül részt vesz az elektrontranszportban (pl. a PS II D1 és D2 fehérjéi), míg mások a stabilitást, a pigmentkötést vagy a szabályozást szolgálják (pl. CP43, CP47 a PS II-ben, PsaA, PsaB a PS I-ben).
A fehérje mátrix kritikusan fontos a reakciócentrum működése szempontjából. A fehérjék hidrofób zsebeket és interakciós pontokat biztosítanak a pigmentek és kofaktorok számára, amelyek így pontosan a megfelelő távolságra és szögben helyezkedhetnek el egymástól. Ez az elrendezés elengedhetetlen a rendkívül gyors és hatékony, szekvenciális elektronátvitelhez, amely a töltésszétválasztás alapja. A fehérjék továbbá védelmet nyújtanak a kofaktoroknak a környezeti hatásokkal szemben, és befolyásolják azok redox potenciálját, optimalizálva az elektronátvitel termodinamikáját.
Pigmentek és kofaktorok precíz elrendezése
A reakciócentrum számos különböző pigmentet és kofaktort tartalmaz, amelyek mindegyike specifikus szerepet játszik az elektronátvitelben. Ezek a molekulák egy szigorúan meghatározott útvonalat alkotnak az elektronok számára:
- Primer donor klorofillok (P): Ezek a speciális klorofill molekulák (gyakran egy dimer, mint a P680 vagy P700) a reakciócentrum szívét képezik. Ők veszik át a fénygyűjtő komplexektől az energiát, és gerjesztett állapotba kerülve adják le első elektronjukat.
- Akcesszórius klorofillok és feofitinek (Chl, Phe): Ezek a molekulák közvetítőként szolgálnak az elektronátvitelben a primer donortól az első stabil akceptor felé. A feofitin különösen érdekes, mivel ez egy klorofill molekula, amelyből hiányzik a központi magnéziumion, és kulcsszerepet játszik a PS II primer akceptoraként.
- Kinonok (QA, QB): Ezek a molekulák stabil elektronakceptorként funkcionálnak, különösen a Type II reakciócentrumokban. Képesek két elektront és két protont felvenni, majd redukált kinol (QH2) formájában továbbadni azokat a fotoszintetikus elektrontranszport lánc következő elemének.
- Vas-kén klaszterek (Fe-S): Különösen a PS I reakciócentrumban és a zöld kén baktériumokban találhatók, ahol kulcsszerepet játszanak az elektronok továbbításában a végső akceptorok felé (pl. FX, FA, FB klaszterek a PS I-ben).
- Karotinoidok: Bár nem vesznek részt közvetlenül az elektronátvitelben, létfontosságú szerepük van a fotoprotekcióban, azáltal, hogy elnyelik a felesleges fényenergiát és semlegesítik a káros reaktív oxigénfajtákat (ROS), megakadályozva ezzel a reakciócentrum károsodását.
Ezeknek a komponenseknek a pontos térbeli elrendezése biztosítja, hogy az elektronátvitel egy rendkívül gyors (pikmásodperces nagyságrendű) és irányított folyamat legyen, minimalizálva az energiaveszteséget és a visszafelé irányuló rekombinációs reakciókat. A finomhangolt távolságok és orientációk optimalizálják a kvantummechanikai alagúthatást, amely kulcsszerepet játszik az elektronok mozgásában.
A töltésszétválasztás mechanizmusa: a fényenergia kémiai energiává alakítása

A reakciócentrum legfontosabb funkciója a töltésszétválasztás, azaz a fényenergia által gerjesztett elektronok elválasztása a primer donortól és stabil akceptorokhoz való továbbítása. Ez a folyamat a fotoszintézis termodinamikai és kinetikai szempontból is legkritikusabb lépése, amely a fényenergiát stabil kémiai energiává alakítja. A töltésszétválasztás rendkívül gyorsan és hatékonyan zajlik le, megakadályozva a gerjesztési energia elvesztését hő vagy fluoreszcencia formájában.
- Fényabszorpció és energiaátvitel: A fénygyűjtő komplexek által elnyelt fényenergia rezonancia transzfer útján, rendkívül gyorsan eljut a reakciócentrum primer donor klorofilljaihoz (pl. P680 a PS II-ben, P700 a PS I-ben). Ez az energiaátadás szinte veszteségmentesen történik.
- Gerjesztés: A primer donor klorofill (P) egy foton elnyelésével vagy energiaátvétellel gerjesztett állapotba (P*) kerül. Ez azt jelenti, hogy az egyik elektronja magasabb energiapályára ugrik, és a molekula átmenetileg instabil, nagy energiájú állapotba kerül.
- Primer töltésszétválasztás: A gerjesztett P* molekula rendkívül gyorsan (néhány pikoszekundum alatt) leadja elektronját egy közeli akceptornak, például egy feofitin molekulának (Phe) a PS II-ben, vagy egy klorofill molekulának a PS I-ben. Ezzel a P molekula oxidálódik (P+), míg az akceptor redukálódik (Phe– vagy Chl–). Ez a legelső és legkritikusabb lépés a fényenergia kémiai energiává alakításában, létrehozva egy töltésszétválasztott állapotot, ahol az elektron és a „lyuk” térben elválasztódik.
- Elektronok stabilizálása: A primer akceptor (pl. Phe–) gyorsan továbbadja az elektront egy stabilabb, másodlagos akceptornak, például egy kinon molekulának (QA). Ez a lépés kulcsfontosságú, mert megakadályozza az elektron visszaáramlását a P+-hoz, és stabilizálja a töltésszétválasztott állapotot (P+QA–). A gyors, szekvenciális elektronátviteli lépések sorozata biztosítja, hogy az energia ne vesszen el hő formájában.
- További elektronátvitel: A QA– továbbadja az elektront egy másik kinon molekulának (QB), amely két elektron felvétele után protonokat is felvesz a sztromából (vagy citoszolból), és redukált kinol (QH2) formájában leválik a reakciócentrumról. Ez a kinol továbbítja az elektronokat a fotoszintetikus elektrontranszport lánc következő elemeinek. A PS I-ben az elektronok vas-kén klasztereken keresztül jutnak el a ferredoxinhoz.
- A primer donor regenerációja: A P+ formában lévő primer donor elektront vesz fel egy másik molekulától, például a vízoxidáló komplexből (PS II esetén) vagy a plasztocianinból (PS I esetén), ezzel visszatér eredeti, redukált állapotába, és készen áll egy újabb foton befogására. Ez a regeneráció biztosítja a reakciócentrum folyamatos működését.
„A reakciócentrum a természet legkifinomultabb napenergia-átalakítója, amely a kvantummechanika elveit kihasználva valósítja meg a fényenergia stabil kémiai energiává alakítását, elindítva az életet fenntartó folyamatokat.”
Ez a folyamat rendkívül hatékony, szinte 100%-os kvantumhatásfokkal megy végbe a primer töltésszétválasztás tekintetében, ami azt jelenti, hogy szinte minden elnyelt foton egy elektronátvitelt indít el. Az elektronok egyirányú áramlása és a gyors stabilizálás kulcsfontosságú a fotoszintézis hatékonyságához és a fotokárosodás minimalizálásához.
A fotoszisztémák: PS I és PS II reakciócentrumok a növényekben
A növényekben, algákban és cianobaktériumokban két fő típusú reakciócentrum működik együtt, szinergikusan, a fényfüggő reakciók során: a Fotoszisztéma II (PS II) és a Fotoszisztéma I (PS I). Ezek szerkezetükben és funkciójukban is eltérnek, de együttműködésük alapvető az oxigéntermelő fotoszintézishez, amely a földi élet alapját képezi.
Fotoszisztéma II (PS II): a vízoxidáció központja
A PS II reakciócentrum egy kinon típusú reakciócentrum, amely szerkezetileg és funkcionálisan hasonlít a purpur baktériumok reakciócentrumához, ami az evolúciós kapcsolatokra utal. Primer donorja a P680, egy speciális klorofill a dimer, amely 680 nm körüli hullámhosszon abszorbeálja a fényt a leghatékonyabban. A PS II a tilakoid membránokban, főként a gránumok stókolt régióiban található meg.
A PS II legfontosabb és legkülönlegesebb funkciója a víz fotolízise, azaz a vízmolekula fényenergia segítségével történő bontása. Ez a folyamat oxigént (O2), protonokat (H+) és elektronokat szabadít fel. A felszabaduló elektronok pótolják a P680 által leadott elektronokat, míg a protonok hozzájárulnak a proton-gradiens kialakításához, amely az ATP szintézishez szükséges. A víz fotolízise a vízoxidáló komplex (OEC – Oxygen Evolving Complex) segítségével történik, amely négy mangánionból, egy kalciumionból és egy kloridionból álló klaszter. Az OEC képes a vízmolekula elektronjait kivonni egy lépésben, négy foton abszorpciója után, egy ciklikus, úgynevezett S-állapotok sorozatán keresztül (S0-S4), amelynek során felhalmozódnak az oxidáló ekvivalensek.
A PS II által leadott elektronok a feofitinen és a QA kinonon keresztül a plasztokinon (PQ) molekulákhoz kerülnek. A QB kinon, amely két elektront és két protont vesz fel, redukált plasztokinol (PQH2) formájában leválik a reakciócentrumról, és a tilakoid membránban diffundálva továbbítja az elektronokat a citokróm b6f komplexhez. Eközben a protonok a sztromából a tilakoid lumenbe pumpálódnak, növelve a proton-gradienst.
A PS II komplex rendkívül érzékeny a fénykárosodásra, különösen a D1 fehérje, amelynek folyamatosan cserélődnie kell a működés fenntartásához. Ez a folyamatos javítási mechanizmus biztosítja a fotoszintézis stabilitását változó fényviszonyok között.
„A Fotoszisztéma II a fotoszintézis motorja, amely nem csupán az elektrontranszportot indítja el, hanem az élet számára nélkülözhetetlen oxigént is termeli a vízmolekula bontásával, ezzel alapvetően formálva bolygónk légkörét és bioszféráját.”
Fotoszisztéma I (PS I): a NADPH termelés központja
A PS I reakciócentrum egy vas-kén típusú reakciócentrum, amely szerkezetileg és funkcionálisan eltér a PS II-től. Primer donorja a P700, egy klorofill a dimer, amely 700 nm körüli hullámhosszon abszorbeálja a fényt a leghatékonyabban. A PS I főként a tilakoid membránok sztromális oldalán, a nem stókolt régiókban található meg.
A PS I elsődleges funkciója a nagy energiájú elektronok továbbítása a ferredoxinhoz (Fd), majd onnan a NADP+ reduktáz enzimhez, amely végül NADP+-ból NADPH-t szintetizál. A NADPH a fényfüggetlen reakciók, a Calvin-ciklus redukáló erejét biztosítja, amelynek során a szén-dioxid cukrokká alakul.
A PS I elektrontranszport lánca a PS II-től kapott elektronokkal indul. A citokróm b6f komplexből származó elektronokat a plasztocianin (PC) szállítja a PS I-hez, ahol azok pótolják a P700 által leadott elektronokat. A P700 gerjesztése után az elektronok egy sor vas-kén klaszteren keresztül jutnak el a ferredoxinhoz. Ezek a vas-kén klaszterek (FX, FA, FB) különböző redox potenciállal rendelkeznek, biztosítva az elektronok egyirányú, downhill áramlását.
A PS I képes ciklikus elektrontranszportot is végezni, amely során az elektronok a ferredoxinból visszaáramlanak a plasztokinon medencébe, majd a citokróm b6f komplexen keresztül ismét a PS I-be. Ez a ciklikus áramlás ATP-t termel, de NADPH-t nem, és fontos szerepet játszik a fotoprotekcióban, valamint a proton-gradiens fenntartásában, különösen magas fényintenzitásnál, amikor a NADPH termelés meghaladja a Calvin-ciklus igényeit.
Az elektrontranszport lánc és az ATP szintézis kapcsolata a reakciócentrumokkal
A reakciócentrumok által elindított elektrontranszport nem öncélú; a célja az ATP (adenozin-trifoszfát) és a NADPH (nikotinamid-adenin-dinukleotid-foszfát) termelése, amelyek a fényfüggetlen reakciók, azaz a Calvin-ciklus energiaforrásai. Az elektronok áramlása a PS II-től a PS I-ig, majd a NADP+ redukciójáig egy komplex láncreakcióban zajlik, amely során protonok pumpálódnak a tilakoid lumenbe, létrehozva egy elektrokémiai gradienset.
Az elektrontranszport lánc főbb állomásai és a reakciócentrumok szerepe:
- Fotoszisztéma II (PS II): Ez a reakciócentrum kezdi az elektrontranszportot a víz fotolízisével. A víz bontása során felszabaduló elektronok pótolják a P680 által leadott elektronokat. A gerjesztett P680 elektronokat ad le a feofitinnek, majd a QA és QB kinonoknak. A QB kinon két elektront és két protont felvéve plasztokinollá (PQH2) redukálódik.
- Plasztokinon (PQ): A PQH2 a membránban diffundálva eljut a citokróm b6f komplexhez. A PQ nem csupán elektronszállító, hanem protonszállító is, amely a sztromából származó protonokat juttatja a tilakoid lumenbe.
- Citokróm b6f komplex: Ez a nagy fehérjekomplex a PQH2-ből származó elektronokat továbbítja a plasztocianinhoz (PC), miközben protonokat pumpál a tilakoid lumenbe. Ez a folyamat az úgynevezett Q-ciklus része, amely hatékonyan növeli a proton-gradiens kialakulását azáltal, hogy egy PQH2 molekula oxidációjával négy protont juttat a lumenbe.
- Plasztocianin (PC): Egy kis, vízoldékony rézproteint tartalmazó molekula, amely az elektronokat szállítja a citokróm b6f komplexből a PS I-hez, áthidalva a két fotoszisztéma közötti távolságot.
- Fotoszisztéma I (PS I): A P700 által leadott elektronokat a PC pótolja. A gerjesztett P700 elektronokat ad le egy sor vas-kén klaszternek (FX, FA, FB), majd a ferredoxinnak (Fd).
- Ferredoxin (Fd): Egy kis, vas-kén proteint tartalmazó molekula, amely az elektronokat szállítja a PS I-ből a ferredoxin-NADP+ reduktáz (FNR) enzimhez.
- Ferredoxin-NADP+ reduktáz (FNR): Ez az enzim két elektront és egy protont felhasználva redukálja a NADP+-ot NADPH-vá. Ez a végső lépés a lineáris elektrontranszport láncban, amely a redukáló erőt szolgáltatja a szén-dioxid fixálásához.
A tilakoid membránon keresztül történő protonpumpálás (a PS II vízoxidációja és a citokróm b6f komplex Q-ciklusa révén) egy jelentős elektrokémiai proton-gradienst hoz létre. A tilakoid lumenben megnő a protonkoncentráció, míg a sztromában csökken. Ez a proton-gradiens egy potenciális energiaforrás, amelyet az ATP-szintáz enzim hasznosít. Az ATP-szintáz egy molekuláris motor, amely a protonok áramlását a lumenből a sztromába (a gradiens mentén) arra használja, hogy ADP-ből és anorganikus foszfátból (Pi) ATP-t szintetizáljon. Ezt a folyamatot fotofoszforilációnak nevezzük, és ez a kémiozmotikus elv alapján működik.
Az ATP és a NADPH ezután a kloroplasztisz sztromájában zajló Calvin-ciklusba kerül, ahol a szén-dioxid fixálódik és cukrokká redukálódik. Így válik a fényenergia stabil kémiai energiává, ami az élet alapját képezi, biztosítva a szerves anyagok szintéziséhez szükséges energiát és redukáló erőt.
A reakciócentrumok evolúciója és diverzitása az élővilágban
A reakciócentrumok nem csupán a modern fotoszintetikus szervezetekben találhatók meg, hanem mélyen gyökereznek az élet evolúciós történelmében. Feltételezések szerint a legkorábbi reakciócentrumok az anoxigén fotoszintetikus baktériumokban jelentek meg, még az oxigéntermelő fotoszintézis kialakulása előtt, ami a Föld légkörét és élővilágát alapjaiban megváltoztatta.
Az anoxigén fotoszintetikus baktériumok reakciócentrumai
Az anoxigén fotoszintézis nem termel oxigént, és a víz helyett más elektron donorokat (pl. H2S, szerves savak, Fe2+) használnak. Két fő típusú reakciócentrumot azonosítottak bennük, amelyek a mai növényi fotoszisztémák őseinek tekinthetők:
- Purpur baktériumok (Type II reakciócentrum): Ezek a baktériumok egy kinon típusú reakciócentrummal rendelkeznek, amely szerkezetileg és funkcionálisan rendkívül hasonlít a növényi PS II-höz. Primer donorjuk a P870 (bakterio-klorofill dimer), amely 870 nm körüli infravörös fényt abszorbeál. Az elektronok ciklikusan áramlanak, ATP-t termelve, de nem termelnek NADPH-t közvetlenül a fényfüggő reakciók során. A redukáló erőt (NADPH) gyakran reverz elektrontranszporttal vagy más mechanizmusokkal állítják elő.
- Zöld kén baktériumok (Type I reakciócentrum): Ezek a baktériumok egy vas-kén típusú reakciócentrummal rendelkeznek, amely funkcionálisan hasonlít a növényi PS I-hez. Primer donorjuk a P840, amely 840 nm körüli infravörös fényt abszorbeál. Képesek NADPH-t termelni, és külső elektron donorokat, mint például a hidrogén-szulfidot (H2S) vagy elemi ként használnak, ami a kéntartalmú vegyületek evolúciós jelentőségére utal.
Ez a két reakciócentrum típus valószínűleg egymástól függetlenül fejlődött ki az ősi Földön, különböző környezeti nyomásokra válaszul. A szerkezeti homológia ellenére a részleteikben eltérő elektronátviteli útvonalakat fejlesztettek ki, alkalmazkodva a rendelkezésre álló elektron donorokhoz és az energiaigényekhez.
Az oxigéntermelő fotoszintézis kialakulása és az endoszimbiotikus elmélet
Az oxigéntermelő fotoszintézis megjelenése, amely a cianobaktériumokban fejlődött ki először mintegy 2,5-3 milliárd évvel ezelőtt, forradalmasította a földi életet. Ez a folyamat a PS II és PS I szinergikus működését igényli, és a vízmolekula elektron donoraként való felhasználásának egyedülálló képességét. A víz bőségesen rendelkezésre álló elektronforrás, ami lehetővé tette a fotoszintézis elterjedését és az oxigén felhalmozódását a légkörben, megnyitva az utat a komplexebb, oxigénfüggő életformák evolúciója előtt, és egyben drasztikusan megváltoztatva a bolygó geokémiáját.
Az endoszimbiotikus elmélet szerint a kloroplasztiszok, a növényi sejtek fotoszintetikus organellumai, ősi cianobaktériumokból származnak, amelyek beépültek egy eukarióta sejtbe. Ez az esemény hozta létre a növényi fotoszintézis alapját, ahol a PS II és PS I reakciócentrumok együttműködve optimalizálták a fényenergia hasznosítását és az oxigéntermelést. Az evolúció során a baktériumok génjeinek nagy része átkerült a gazdasejt magjába, de a reakciócentrumok és más kulcsfontosságú fotoszintetikus komponensek továbbra is a kloroplasztiszok genomjában kódolódnak, bizonyítva ősi eredetüket.
Ez az evolúciós történet rávilágít a reakciócentrumok alapvető fontosságára nemcsak az egyedi szervezetek életében, hanem a bolygó bioszférájának globális fejlődésében is. A reakciócentrumok kialakulása és finomhangolása kulcsfontosságú lépés volt az élet történetében, amely lehetővé tette a komplex ökoszisztémák kialakulását és a földi környezet radikális átalakulását, megalapozva a ma ismert biológiai sokféleséget.
A reakciócentrumok szabályozása és védelme a fotokárosodástól

A reakciócentrumok rendkívül hatékonyak a fényenergia átalakításában, de egyben érzékenyek is a környezeti stresszre, különösen az intenzív fényre. A túlzott fényenergia károsíthatja a komplexet, különösen a PS II-t, ami fotokárosodáshoz és a fotoszintézis hatékonyságának csökkenéséhez vezethet. Ezért a növények és algák kifinomult mechanizmusokat fejlesztettek ki a reakciócentrumok védelmére és működésük szabályozására, biztosítva a túlélést a változó fényviszonyok között.
Fotoprotekciós mechanizmusok a reakciócentrumok védelmében
A fotoprotekció célja a felesleges fényenergia elvezetése és a káros reaktív oxigénfajták (ROS) képződésének megakadályozása, amelyek a túlzott gerjesztési energia hatására keletkezhetnek. Néhány kulcsfontosságú mechanizmus:
- Nem-fotokémiai kioltás (NPQ – Non-Photochemical Quenching): Ez egy gyors és reverzibilis folyamat, amely során a felesleges fényenergia hővé alakul és leadódik, még mielőtt elérné a reakciócentrumot. Az NPQ mechanizmus a fénygyűjtő komplexekben zajlik, és a tilakoid lumen pH-jának változásai aktiválják. A karotinoidok (különösen a xantofillok, mint a zeaxantin) és specifikus fehérjék (pl. PsbS a PS II-ben) kulcsszerepet játszanak ebben a disszipációs folyamatban, megváltoztatva az antenna komplex konformációját.
- Karotinoidok szerepe: A karotinoidok nemcsak antennapigmentként funkcionálnak, hanem közvetlenül is részt vesznek a fotoprotekcióban. Képesek elnyelni az energiát a gerjesztett klorofilloktól, megakadályozva a szingulett oxigén képződését, amely rendkívül reaktív és károsíthatja a membránokat és fehérjéket. Emellett semlegesítik a már kialakult reaktív oxigénfajtákat is, mint antioxidánsok.
- Ciklikus elektrontranszport: Különösen a PS I-nél megfigyelhető, hogy az elektronok a ferredoxinból visszakerülnek a plasztokinon medencébe, majd a citokróm b6f komplexen keresztül ismét a PS I-be. Ez a ciklikus áramlás ATP-t termel, de NADPH-t nem, és segíthet a felesleges fényenergia elvezetésében, valamint a proton-gradiens fenntartásában, ami az ATP szintézishez szükséges. Ezáltal csökkenti a PS I túlredukáltságát és a ROS képződés kockázatát.
A D1 fehérje turnover a PS II-ben: egy dinamikus javítási mechanizmus
A PS II reakciócentrum különösen érzékeny a fénykárosodásra, főként a D1 fehérje. Ez a fehérje a PS II magjának része, és az elektronátvitelben kulcsszerepet játszó pigmenteket és kofaktorokat köti meg. Erős fény hatására a D1 fehérje irreverzibilisen károsodhat (pl. oxidatív károsodás vagy a fehérje szerkezetének denaturációja révén), ami a PS II működésének romlásához vezet. A növények azonban rendkívül hatékony és folyamatos javító mechanizmussal rendelkeznek, amely biztosítja a rendszer rugalmasságát:
- A károsodott D1 fehérjét specifikus proteázok (pl. FtsH proteázok) gyorsan felismerik és lebontják.
- Egy új D1 fehérje de novo szintetizálódik a kloroplasztisz riboszómáin, és beépül a PS II komplexbe a membránban.
- A PS II komplex újra aktiválódik és működőképes lesz.
Ez a folyamatos lebontás és szintézis, az úgynevezett D1 fehérje turnover, biztosítja a PS II folyamatos működését még stresszes körülmények között is. A turnover sebessége nagymértékben függ a fényintenzitástól; minél erősebb a fény, annál gyorsabban zajlik a D1 fehérje cseréje. Ez a dinamikus alkalmazkodás létfontosságú a növények túléléséhez a változó fényviszonyok között, és egyben rávilágít a reakciócentrumok komplex szabályozására.
A reakciócentrumok védelmére szolgáló mechanizmusok rendkívül összetettek és szinergikusan működnek, biztosítva a fotoszintézis stabilitását és hatékonyságát a környezeti kihívások ellenére. Ezen mechanizmusok mélyebb megértése kulcsfontosságú a mezőgazdasági termelékenység növeléséhez és a növények stressztűrő képességének javításához.
A reakciócentrumok vizsgálata: módszerek és történelmi felfedezések
A reakciócentrumok szerkezetének és működésének megértése évtizedes, intenzív kutatások eredménye, amelyek során számos fejlett biokémiai és biofizikai módszert alkalmaztak. Ezek a módszerek lehetővé tették, hogy atomi szinten vizsgálhassuk meg ezen komplexek működését, és feltárjuk a fényenergia átalakításának alapvető elveit.
Röntgenkrisztallográfia: az atomi szerkezet feltárása
Az egyik legjelentősebb áttörést a röntgenkrisztallográfia hozta el a reakciócentrumok kutatásában. 1985-ben Johann Deisenhofer, Hartmut Michel és Robert Huber meghatározta a purpur baktérium (Rhodopseudomonas viridis) reakciócentrumának atomi szerkezetét. Ez a felfedezés, amelyért 1988-ban kémiai Nobel-díjat kaptak, először mutatta meg részletesen, hogyan helyezkednek el a fehérjék és a kofaktorok a membránban, és hogyan valósul meg az elektronátvitel. Ez a munka alapot szolgáltatott az összes többi reakciócentrum, beleértve a növényi PS II és PS I megértéséhez is, mivel a purpur baktériumok reakciócentruma szerkezeti homológiát mutat a PS II-vel.
A röntgenkrisztallográfia segítségével kapott atomi felbontású képek kulcsfontosságúak voltak az elektronátvitel útvonalainak, a pigmentek elrendezésének és a protein-kofaktor kölcsönhatások megértésében. Ezek az ismeretek nemcsak a fotoszintézis alapjait világították meg, hanem inspirációt adtak a bioinspirált energiaátalakító rendszerek tervezéséhez is, lehetővé téve a természetes folyamatok lemásolását mesterséges rendszerekben.
Spektroszkópiai módszerek a dinamikus folyamatok vizsgálatára
A reakciócentrumok dinamikus folyamatainak, különösen az elektronátvitel rendkívül gyors lépéseinek vizsgálatára különböző spektroszkópiai módszereket alkalmaznak, amelyek lehetővé teszik a molekulák fényelnyelési és emissziós tulajdonságainak elemzését:
- Abszorpciós spektroszkópia: Lehetővé teszi a pigmentek fényelnyelési tulajdonságainak vizsgálatát és a reakciócentrum primer donor klorofilljainak azonosítását (pl. P680, P700), azáltal, hogy méri a fényelnyelés spektrumát.
- Fluoreszcencia spektroszkópia: A fotoszintézis hatékonyságának és a fotokémiai reakciók sebességének mérésére használják. A klorofill fluoreszcencia intenzitása és kinetikája közvetlenül összefügg a reakciócentrumok állapotával és az elektronátvitel hatékonyságával.
- Femtoszekundumos tranziens abszorpciós spektroszkópia: Ez a legfejlettebb módszer lehetővé teszi az elektronátvitel ultragyors (pikmásodperces, femtoszekundumos) lépéseinek valós idejű követését. Segítségével feltárták a töltésszétválasztás pontos kinetikáját és az intermedier állapotokat, amelyek a fényelnyeléstől a stabil töltésszétválasztásig vezetnek.
- Elektron spin rezonancia (ESR) spektroszkópia: A paramágneses intermedierek (pl. P+, QA–) azonosítására és szerkezetük vizsgálatára alkalmas, amelyek az elektronátvitel során keletkeznek. Az ESR segítségével meghatározható az elektronok lokalizációja és a kofaktorok közötti távolság.
- Krio-elektronmikroszkópia (Cryo-EM): Az utóbbi években a cryo-EM forradalmasította a nagy méretű fehérjekomplexek szerkezetvizsgálatát, lehetővé téve a reakciócentrumok és fénygyűjtő komplexek térbeli szerkezetének meghatározását közel atomi felbontással, anélkül, hogy kristályt kellene növeszteni.
Genetikai és biokémiai megközelítések
A genetikai manipuláció, mint például a pontmutagenezis vagy génkiütés, lehetővé teszi a reakciócentrum egyes alegységeinek vagy specifikus aminosav-maradékainak funkciójának vizsgálatát. A mutáns törzsek tanulmányozása révén a kutatók azonosíthatják azokat a kulcsfontosságú aminosavakat, amelyek az elektronátvitelben, a pigmentkötésben vagy a komplex stabilitásában játszanak szerepet. A biokémiai tisztítási és rekonsztitúciós technikák révén a kutatók izolálhatják a reakciócentrumokat, és in vitro körülmények között tanulmányozhatják működésüket, meghatározva a különböző komponensek szerepét és kölcsönhatásait.
Ezeknek a módszereknek a kombinációja adja a reakciócentrumokról szerzett rendkívül részletes tudásunkat, a molekuláris szintű szerkezetüktől a dinamikus funkciójukig. Az eredmények nemcsak az alapvető biológiai folyamatok megértését mozdítják elő, hanem új utakat nyitnak meg a bioenergetikai technológiák és a szintetikus biológia fejlesztése felé is.
A reakciócentrum jelentősége a bioenergetikában és a biotechnológiában
A reakciócentrumok nem csupán elméleti érdekességek; működésük alapvető betekintést nyújt a rendkívül hatékony energiaátalakítás elveibe, ami inspirációt jelent a modern tudomány és technológia számára. A reakciócentrumok tanulmányozása és megértése kulcsfontosságú a bioenergetika, a biotechnológia és a fenntartható energiatermelés jövője szempontjából, mivel ezek a természetes rendszerek példát mutatnak a hatékony napenergia-hasznosításra.
Mesterséges fotoszintézis és napenergia hasznosítás
Az egyik legizgalmasabb kutatási terület a mesterséges fotoszintézis, amelynek célja a természetes fotoszintézis elveinek alkalmazása szintetikus rendszerekben a napenergia hatékony kémiai energiává (pl. hidrogén üzemanyaggá, szerves molekulákká) történő átalakítására. A reakciócentrumok mintájára fejlesztett mesterséges rendszerek képesek lehetnek a fény befogására és töltésszétválasztásra, utánozva a természetes folyamatok hatékonyságát és stabilitását.
A kutatók olyan molekuláris szerkezeteket terveznek, amelyek utánozzák a primer donor-akceptor párok precíz elrendeződését, és képesek stabilan fenntartani a töltésszétválasztott állapotot. Ez magában foglalja a megfelelő pigmentek (pl. porfirin származékok), elektron akceptorok (pl. kinonok, fullerének) és donorok (pl. vízoxidáló katalizátorok) kiválasztását és precíz elrendezését mesterséges fehérje vagy polimer vázakban. A kihívás abban rejlik, hogy olyan szintetikus rendszereket hozzanak létre, amelyek nemcsak hatékonyak, hanem stabilak és gazdaságosan gyárthatók is.
A mesterséges fotoszintézis ígéretes utat kínál a tiszta, megújuló energiaforrások fejlesztéséhez, amelyek képesek lehetnek a napfényből üzemanyagot termelni, ezzel csökkentve a fosszilis energiahordozóktól való függőséget és enyhítve az éghajlatváltozást. Ez a technológia hosszú távon forradalmasíthatja az energiatermelést, és hozzájárulhat egy fenntarthatóbb jövőhöz.
Biotechnológiai alkalmazások és herbicidek célpontjai
A reakciócentrumok, különösen a PS II, célpontként szolgálnak számos herbicidek (gyomirtó szerek) számára. Ezek a vegyületek gyakran a PS II elektrontranszport láncát gátolják, például a QB kötőhelyén versengve a kinonnal, vagy közvetlenül károsítva a D1 fehérjét. Azáltal, hogy blokkolják az elektrontranszportot, megakadályozzák az ATP és NADPH termelését, ami a gyomnövények elpusztulásához vezet. A reakciócentrumok részletes szerkezetének ismerete segíti a szelektívebb és hatékonyabb herbicidek tervezését, minimalizálva a környezeti mellékhatásokat és a rezisztencia kialakulásának kockázatát.
Ezen túlmenően, a reakciócentrumok génjeinek módosítása vagy optimalizálása révén a biotechnológia potenciálisan növelheti a terménynövények fotoszintetikus hatékonyságát. Ez magában foglalhatja az antenna komplexek méretének módosítását (pl. kisebb antennákkal csökkentve a fényveszteséget sűrű állományban), a reakciócentrumok fényadaptációs képességének javítását, vagy a D1 fehérje turnover sebességének optimalizálását, ami hozzájárulhat a globális élelmiszerbiztonság javításához a növekvő népesség és a változó éghajlat kihívásai közepette.
A reakciócentrumok mint nanotechnológiai építőkövek és bioszenzorok
A reakciócentrumok kivételes stabilitásuk és precíz molekuláris architektúrájuk miatt potenciális nanotechnológiai építőkövekként is szolgálhatnak. Kutatások folynak arról, hogyan lehetne izolált reakciócentrumokat vagy azok szintetikus analógjait beépíteni nanostrukturált anyagokba vagy eszközökbe, például bioszenzorokba, nanoelektronikai komponensekbe vagy biokatalizátorokba. Az ilyen biohibrid rendszerek kihasználnák a reakciócentrumok természetes fényérzékenységét és töltésszétválasztó képességét, új funkcionális anyagok és alkalmazások létrehozására, például a környezeti szennyező anyagok kimutatására vagy a gyógyszerkutatásban.
A reakciócentrumok tanulmányozása tehát nem csupán az alapvető biológiai folyamatok megértéséhez járul hozzá, hanem gyakorlati alkalmazások széles skáláját is megnyitja, a fenntartható energiatermeléstől az agrártudományokig és a nanotechnológiáig. A természetes rendszerekből nyert inspiráció kulcsfontosságú a jövő technológiai kihívásainak megoldásában, és új utakat nyit meg az emberiség számára.
Összefoglaló táblázat: A Fotoszisztéma II és I reakciócentrumok összehasonlítása
Az alábbi táblázatban összefoglaltuk a két fő típusú reakciócentrum, a Fotoszisztéma II és I legfontosabb jellemzőit, hogy áttekinthetőbbé tegyük a köztük lévő különbségeket és hasonlóságokat, amelyek a fotoszintézis hatékony működését biztosítják.
| Jellemző | Fotoszisztéma II (PS II) | Fotoszisztéma I (PS I) |
|---|---|---|
| Primer Donor | P680 (klorofill a dimer) | P700 (klorofill a dimer) |
| Fényabszorpciós Maximum | ~680 nm | ~700 nm |
| Reakciócentrum Típusa | Kinon típusú (Type II) | Vas-kén típusú (Type I) |
| Elektron Donor | Víz (H2O) az OEC-n keresztül | Plasztocianin (PC) |
| Elektron Akceptor | Plasztokinon (PQ) | Ferredoxin (Fd) |
| Fő Funkció | Víz fotolízise, oxigéntermelés, protonpumpálás a lumenbe | NADP+ redukciója NADPH-vá, protonpumpálás (ciklikus ETS esetén) |
| Termékek | O2, H+, elektronok a PQ-nak | Elektronok az Fd-nek, NADPH (az FNR-en keresztül), ATP (ciklikus ETS esetén) |
| Sérülékenység | Nagyon érzékeny a fénykárosodásra (D1 fehérje turnover) | Kevésbé érzékeny, de részt vesz a ciklikus ETS-ben a fotoprotekcióban |
| Elhelyezkedés a tilakoid membránban | Főként a gránumok stókolt régióiban | Főként a sztromális tilakoidokban és a gránumok szélén |
| Evolúciós Homológia | Purpur baktériumok reakciócentruma | Zöld kén baktériumok reakciócentruma |
Ez a táblázat egyértelműen bemutatja, hogy bár mindkét fotoszisztéma a reakciócentrum elvét használja a fényenergia átalakítására, specifikus feladataik és mechanizmusuk eltérő, kiegészítve egymást az oxigéntermelő fotoszintézisben. Együttműködésük teszi lehetővé a fényenergia maximális hasznosítását és a stabil kémiai energia előállítását.
Jövőbeli kutatási irányok és a reakciócentrumokkal kapcsolatos kihívások

Annak ellenére, hogy a reakciócentrumokról már rendkívül sok információ áll rendelkezésre, számos nyitott kérdés és kihívás vár még megoldásra a tudományos közösség előtt. A jövőbeli kutatások célja nem csupán a még mélyebb megértés, hanem a tudás gyakorlati alkalmazása is a globális problémák kezelésére, mint például az élelmiszerbiztonság és a tiszta energia.
Kvantummechanikai jelenségek a fotoszintézisben
Az utóbbi években egyre nagyobb figyelem irányul a kvantummechanikai jelenségekre a fotoszintézisben. Feltételezések szerint az energiaátvitel a fénygyűjtő komplexekben és talán még a reakciócentrum primer töltésszétválasztásában is kvantum koherencia révén zajlik, ami magyarázatot adhat a rendkívüli hatékonyságra és a szinte veszteségmentes energiaátvitelre. A kvantum biológia ezen területének vizsgálata új elméleti kereteket és kísérleti módszereket igényel, és alapjaiban változtathatja meg a fényenergia átalakításáról alkotott képünket, új utakat nyitva meg a kvantumtechnológiák fejlesztésében.
A fotoszintézis hatékonyságának növelése a mezőgazdaságban
A globális népességnövekedés és az éghajlatváltozás kihívásai rávilágítanak arra, hogy sürgősen növelni kell a terménynövények fotoszintetikus hatékonyságát. A reakciócentrumok, mint a fényenergia hasznosításának kulcsfontosságú pontjai, potenciális célpontok a genetikai módosítások számára. A kutatók azt vizsgálják, hogyan lehetne optimalizálni a fénygyűjtő komplexek méretét (pl. kisebb antennákkal, amelyek kevésbé telítődnek túl erős fényben), a reakciócentrumok fényadaptációs képességét, vagy akár a D1 fehérje turnover sebességét, hogy a növények jobban alkalmazkodjanak a változó fényviszonyokhoz és hatékonyabban hasznosítsák a napfényt. Ez hozzájárulhat a terméshozam növeléséhez és a fenntartható mezőgazdaság kialakításához.
Biohibrid rendszerek és új anyagok fejlesztése
A reakciócentrumok, vagy azok kulcsfontosságú elemeinek integrálása mesterséges anyagokba, úgynevezett biohibrid rendszerek létrehozását teszi lehetővé. Ezek az anyagok a biológiai rendszerek önszerveződő képességét és hatékonyságát ötvöznék a mesterséges anyagok stabilitásával és tervezhetőségével. Ilyen rendszerek felhasználhatók lehetnek új generációs napelemek, biológiai szenzorok vagy akár üzemanyagcellák fejlesztésére. A cél olyan rendszerek létrehozása, amelyek tartósak, költséghatékonyak és nagy volumenben gyárthatók, miközben megőrzik a természetes reakciócentrumok kivételes hatékonyságát.
A fotoszintézis sötét oldala: a fotokárosodás és stressztűrés
Bár a növények rendelkeznek kifinomult fotoprotekciós mechanizmusokkal, a túlzott fényenergia továbbra is komoly problémát jelenthet, különösen a mezőgazdaságban. A reakciócentrumok károsodásának mélyebb megértése és a javító mechanizmusok finomhangolása új stratégiákat kínálhat a termésveszteségek csökkentésére és a növények stressztűrő képességének növelésére. A D1 fehérje turnover mechanizmusának részletesebb feltárása például hozzájárulhat a rezisztens növényfajták nemesítéséhez, amelyek jobban ellenállnak a környezeti stressznek, mint például a szárazságnak vagy a magas hőmérsékletnek, amelyek fokozzák a fénykárosodást.
A reakciócentrumok kutatása tehát továbbra is a biológia, a kémia és a fizika határterületén zajló, dinamikus és izgalmas terület. Az ezen a területen elért eredmények nemcsak az élet alapvető mechanizmusainak megértéséhez járulnak hozzá, hanem konkrét megoldásokat is kínálnak a modern társadalom energia- és környezeti kihívásaira, egy fenntarthatóbb és élhetőbb jövő felé mutatva.
A reakciócentrum, mint a fotoszintézis központi eleme, egy olyan molekuláris csoda, amely évmilliárdok alatt fejlődött ki a tökéletes hatékonyság elérésére. Jelentőségét nem lehet eléggé hangsúlyozni, hiszen ez a parányi gépezet felelős azért a folyamatért, amely a napfényt az élet üzemanyagává alakítja, fenntartva bolygónk ökoszisztémáját és biztosítva az oxigént, amit belélegzünk. A róla szerzett tudásunk továbbra is inspirálja a tudósokat és mérnököket, hogy új utakat találjanak a fenntartható jövő megteremtésére, a természet bölcsességét felhasználva a tudományos innovációban.







