
A sejtek energiaellátása az élet egyik legalapvetőbb és legkomplexebb folyamata, melynek középpontjában az adenozin-trifoszfát, röviden ATP, áll. Az ATP az univerzális energiatároló molekula, amely a sejtek összes anyagcsere-folyamatához, mozgásához, szállításához és szintéziséhez biztosítja a szükséges energiát. Ezt az életfontosságú molekulát a sejt két fő útvonalon szintetizálja: a szubsztrát-szintű foszforiláció és az oxidatív foszforiláció révén. Míg az előbbi kisebb mennyiségű ATP-t termel közvetlenül bizonyos anyagcsere-reakciók során, addig az oxidatív foszforiláció a sejtlégzés aerob szakaszának messze legproduktívabb része, amely a sejt energiaigényének döntő többségét fedezi.
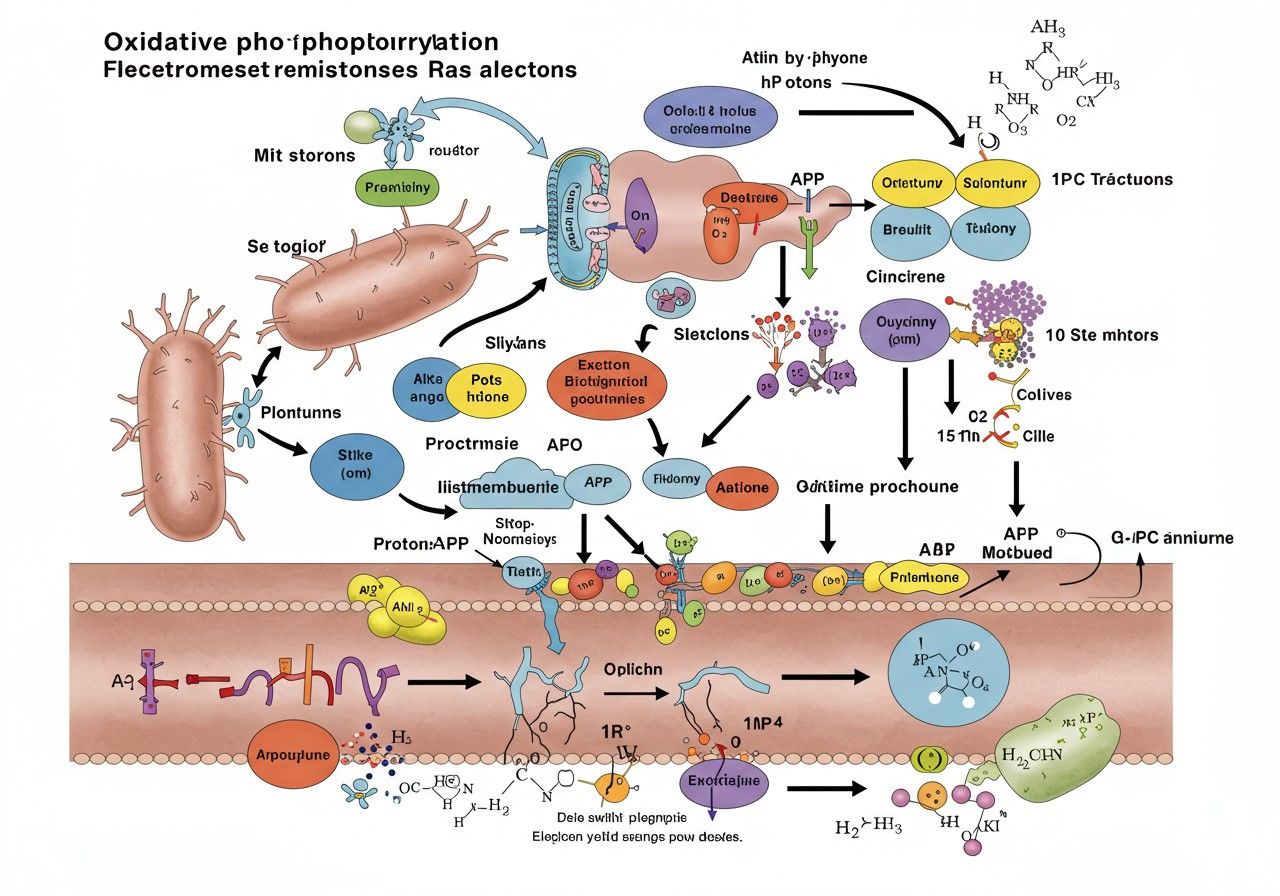
Az oxidatív foszforiláció egy rendkívül elegáns és hatékony biokémiai mechanizmus, amely a legtöbb eukarióta sejtben a mitokondriumokban zajlik. Lényege, hogy a tápanyagok lebontásából származó elektronok energiáját felhasználva proton gradienst hoz létre egy membránon keresztül, majd ezt a gradienst használja fel az ATP szintézisére. Ez a folyamat két szorosan összekapcsolódó részből áll: az elektron transzport láncból (ETL) és a kémiozmotikus kapcsolásból, melyet az ATP szintáz enzim hajt végre. Ennek a bonyolult gépezetnek a megértése kulcsfontosságú az élet alapvető működésének felfogásához, és számos betegség patomechanizmusának magyarázatához.
Mi az oxidatív foszforiláció lényege?
Az oxidatív foszforiláció egy olyan anyagcsere-útvonal, amelyben a sejtek a tápanyagok, például glükóz vagy zsírsavak, oxidációjából felszabaduló energiát hasznosítják ATP előállítására. A folyamat neve is utal a két kulcsfontosságú komponensre: az „oxidatív” rész az elektronok oxidációjára és a membránon keresztüli transzportjára vonatkozik, míg a „foszforiláció” az ADP (adenozin-difoszfát) foszforilációjára, azaz egy foszfátcsoport hozzáadására utal, aminek eredményeként ATP keletkezik. Ez a mechanizmus a legfőbb ATP-termelő útvonal az aerob szervezetekben, amelyek oxigént használnak fel a tápanyagok lebontásához.
A folyamat során a tápanyagok lebontásából származó elektronok, melyeket NADH és FADH₂ molekulák szállítanak, egy sor fehérjekomplexen keresztül áramlanak a mitokondrium belső membránjában. Ez az elektronáramlás energiát szabadít fel, amelyet arra használnak, hogy protonokat (H⁺ ionokat) pumpáljanak a mitokondrium mátrixából a membránok közötti térbe. Ezzel létrejön egy elektrokémiai proton gradiens, amely potenciális energiát tárol. Ezt a potenciális energiát használja fel aztán az ATP szintáz enzim, hogy a protonok visszaáramlását felhasználva ATP-t szintetizáljon az ADP-ből és anorganikus foszfátból (Pi).
Az oxidatív foszforiláció a sejt energiaellátásának mesterműve, ahol az elektronok tánca és a protonok áramlása teremti meg az élethez szükséges univerzális energiamolekulát, az ATP-t.
Ez a szinergikus működés teszi lehetővé, hogy egyetlen glükózmolekula teljes oxidációjából akár 30-32 molekula ATP is keletkezzen, szemben a glikolízis során keletkező mindössze 2 ATP-vel. Az oxidatív foszforiláció tehát a sejtek energiaháztartásának motorja, amely nélkül a komplexebb életformák létezése elképzelhetetlen lenne.
A mitokondrium: az oxidatív foszforiláció színtere
Az oxidatív foszforiláció folyamata szinte kizárólag a mitokondriumokban, az eukarióta sejtek „energiagyáraiban” zajlik. Ezek az organellumok különleges szerkezetükkel optimális környezetet biztosítanak az elektron transzport lánc és az ATP szintézis számára. A mitokondriumoknak két membránjuk van: egy külső és egy belső membrán. A külső mitokondriális membrán viszonylag áteresztő, számos porin nevű fehérjét tartalmaz, amelyek lehetővé teszik kisebb molekulák átjutását a citoplazmából a membránok közötti térbe.
A belső mitokondriális membrán ezzel szemben erősen szelektíven áteresztő, és számos redővel, úgynevezett krisztával rendelkezik, amelyek jelentősen megnövelik a membrán felületét. Ez a nagy felület kulcsfontosságú, mivel a belső membránon helyezkednek el az elektron transzport lánc fehérjekomplexei és az ATP szintáz. A membránok közötti teret és a belső membrán által körülhatárolt belső teret, a mátrixot, a belső mitokondriális membrán választja el. A mátrixban zajlik a citrátkör (Krebs-ciklus) és a zsírsavak β-oxidációja, amelyek a NADH és FADH₂ molekulákat termelik, azaz az elektron transzport lánc „üzemanyagát”.
A mitokondriális szerkezet tehát tökéletesen adaptálódott az oxidatív foszforiláció hatékony végrehajtására. A belső membrán redőzöttsége maximalizálja az enzimek és transzportfehérjék számát, míg a két membrán közötti tér és a mátrix közötti szigorú elválasztás lehetővé teszi a proton gradiens kialakítását és fenntartását. A mitokondriumok emellett saját DNS-sel és riboszómákkal is rendelkeznek, ami arra utal, hogy endoszimbiózis révén kerülhettek be az eukarióta sejtekbe, és önállóan képesek bizonyos fehérjéket szintetizálni a saját működésükhöz.
Az elektron transzport lánc (ETL): az energia „betakarítása”
Az oxidatív foszforiláció elsődleges lépése az elektron transzport lánc (ETL), más néven légzési lánc. Ez egy sor, a mitokondrium belső membránjában elhelyezkedő fehérjekomplexből áll, amelyek szekvenciálisan továbbítják az elektronokat. Az elektronok forrása a glikolízis, a piruvát oxidáció és a citrátkör során keletkező redukált koenzimek, elsősorban a NADH (nikotinamid-adenin-dinukleotid) és a FADH₂ (flavin-adenin-dinukleotid). Ezek a molekulák nagy energiájú elektronokat szállítanak az ETL-hez, ahol azok fokozatosan, lépésről lépésre, egyre nagyobb elektronegativitású akceptorokhoz vándorolnak.
Az elektronok áramlása során felszabaduló energia nem vész el hő formájában (legalábbis nem teljesen), hanem arra használódik, hogy protonokat (H⁺) pumpáljon a mitokondrium mátrixából a membránok közötti térbe. Ez a folyamat elektromos és kémiai potenciálkülönbséget hoz létre a membrán két oldala között, amelyet proton-mozgató erőnek (PMF) neveznek. A protonok pumpálását három fő fehérjekomplex végzi az ETL-ben: az I., III. és IV. komplex.
Az elektron transzport lánc komplexek
Az ETL öt fő komplexből áll, amelyek közül négy (I, II, III, IV) részt vesz az elektronok szállításában, az ötödik (V) pedig az ATP szintézisében.
I. komplex: NADH-dehidrogenáz
Ez a legnagyobb komplex, amely a NADH-tól fogadja az elektronokat. A NADH leadja két elektronját a komplexnek, oxidálódva NAD⁺-t képez. Az elektronok ezután a komplexen belül található FMN (flavin-mononukleotid) és számos vas-kén centrum segítségével jutnak tovább. Az elektronok áramlása során a I. komplex négy protont pumpál a mátrixból a membránok közötti térbe.
II. komplex: Szukcinát-dehidrogenáz
Ez a komplex egyedülálló, mivel egyúttal a citrátkör egyik enzime is. Közvetlenül a FADH₂-től fogadja az elektronokat, amely a szukcinát fumaráttá történő oxidációja során keletkezik. A FADH₂ szintén két elektront ad le, oxidálódva FAD-dá. Fontos különbség, hogy a II. komplex nem pumpál protonokat a membránon keresztül. Az elektronok a FADH₂-től vas-kén centrumokon keresztül jutnak tovább.
Ubikinon (Q) – a mobil elektronhordozó
Az I. és II. komplexekből származó elektronok egy hidrofób, lipidoldékony molekulához, az ubikinonhoz (koenzim Q10) jutnak. Az ubikinon mobilis hordozóként funkcionál a belső membránban, és az I. és II. komplexből a III. komplexhez szállítja az elektronokat. Az ubikinon képes két elektront és két protont felvenni, redukálódva ubikinollá (QH₂), majd leadva azokat, oxidálódva ubikinonná.
III. komplex: Citokróm c reduktáz (Citokróm bc₁ komplex)
A III. komplex az ubikinoltól fogadja az elektronokat. Ez a komplex több citokrómot (vasat tartalmazó hem csoporttal rendelkező fehérjék) és vas-kén centrumokat tartalmaz. A III. komplex a Q-ciklus nevű mechanizmuson keresztül továbbítja az elektronokat, amelynek során két protont pumpál a mátrixból a membránok közötti térbe minden két elektron továbbítása során.
Citokróm c – egy másik mobil elektronhordozó
A III. komplexből az elektronok egy másik mobil hordozóhoz, a citokróm c-hez jutnak. A citokróm c egy kis, vízoldékony fehérje, amely a mitokondrium membránok közötti terében helyezkedik el. Egyetlen elektront képes szállítani a III. komplexből a IV. komplexbe.
IV. komplex: Citokróm c oxidáz
Ez az utolsó komplex a láncban, amely a citokróm c-től fogadja az elektronokat. Ez a komplex tartalmaz réz centrumokat és citokróm a és a₃-at. A IV. komplex rendkívül fontos, mivel itt történik meg az elektronok végső akceptorához, az oxigénhez (O₂) történő átadása. Az oxigén felveszi az elektronokat és protonokat a mátrixból, vízzé (H₂O) redukálódik. Ez a lépés teszi az oxidatív foszforilációt aerob folyamattá. A IV. komplex két protont pumpál a membránok közötti térbe minden két elektron továbbítása és egy fél molekula oxigén redukciója során.
Az elektron transzport lánc olyan, mint egy biológiai vízesés: az elektronok lépcsőzetes esése során felszabaduló energia hajtja a protonok „felpumpálását”, létrehozva azt a potenciális energiát, amely az ATP szintéziséhez szükséges.
Az elektronok áramlása az ETL-ben egyirányú, a nagy energiájú NADH/FADH₂-től az alacsony energiájú oxigénig. Az energiafelszabadulás fokozatos, ami maximalizálja az ATP-szintézis hatékonyságát és minimalizálja az energiaveszteséget hő formájában. Az oxigén hiánya blokkolja a IV. komplex működését, ami az egész ETL leállásához és így az ATP termelés drasztikus csökkenéséhez vezet, ami gyorsan halálos lehet a sejt számára.
Kémiozmotikus kapcsolás: a proton gradiens és az ATP szintáz
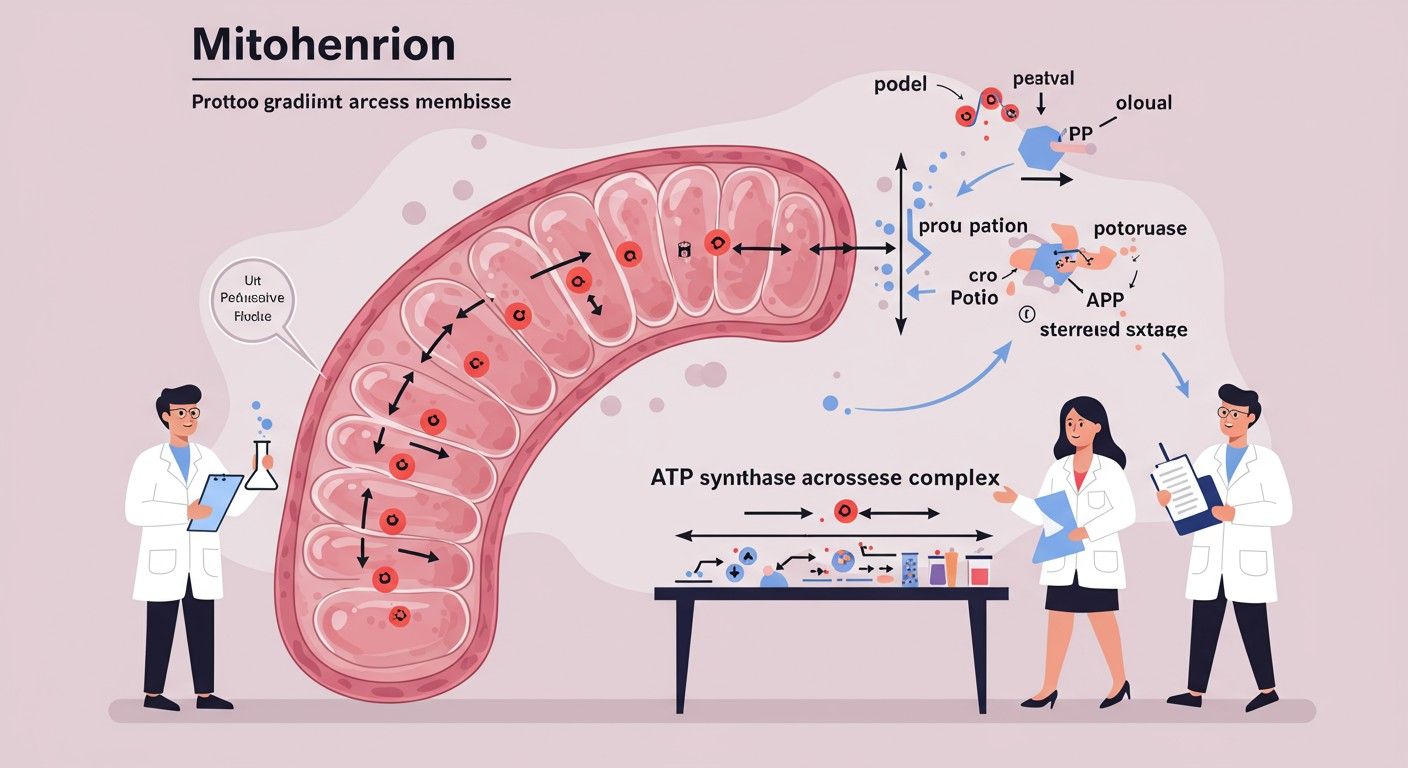
Az elektron transzport lánc működésének közvetlen következménye a proton gradiens kialakulása a mitokondrium belső membránja mentén. A protonok (H⁺ ionok) pumpálása a mátrixból a membránok közötti térbe kétféle potenciálkülönbséget hoz létre:
- Kémiai potenciálkülönbség (pH gradiens): A membránok közötti térben megnő a protonkoncentráció, így a pH csökken, míg a mátrixban a protonkoncentráció csökken, így a pH nő.
- Elektromos potenciálkülönbség (membránpotenciál): A membránok közötti tér pozitívabb töltésűvé válik a felhalmozódott protonok miatt, míg a mátrix negatívabb töltésű marad.
Ezt a két komponenst együttesen elektrokémiai proton-gradiensnek vagy proton-mozgató erőnek (PMF) nevezzük. Ez a gradiens egyfajta „energiaszámla”, amely potenciális energiát tárol, hasonlóan egy felduzzasztott víztározóhoz. A protonok természetüknél fogva igyekeznek visszajutni a mátrixba, ahol alacsonyabb a protonkoncentráció és negatívabb a töltés, de a belső mitokondriális membrán számukra nagyrészt átjárhatatlan.
Az ATP szintáz működése
A protonok visszaáramlását egy speciális enzimkomplex, az ATP szintáz (V. komplex) teszi lehetővé, amely egyben felhasználja a protonok áramlásából felszabaduló energiát az ATP szintézisére. Az ATP szintáz egy molekuláris motor, amely két fő részből áll:
- F₀ alegység: Ez a rész a belső mitokondriális membránba ágyazódik be, és egy protoncsatornaként funkcionál. A F₀ alegység alkotja azt a „rotort”, amely a protonok áthaladása során forog.
- F₁ alegység: Ez a rész a mitokondrium mátrixában helyezkedik el, és az ATP szintézis katalitikus helyét tartalmazza. Az F₁ alegység áll egy „állórészből” és egy „rotorból”, amely a F₀ forgását követi.
Amikor a protonok a membránok közötti térből a F₀ alegységen keresztül visszaáramlanak a mátrixba a koncentráció- és töltésgradiens mentén, a F₀ alegység forogni kezd. Ez a forgás mechanikai energiát ad át az F₁ alegységnek, amely konformációs változásokat idéz elő a katalitikus helyein. Ezek a konformációs változások teszik lehetővé az ADP és a Pi (anorganikus foszfát) kötődését, majd az ATP kialakulását. Az F₁ alegység minden 360 fokos fordulata három ATP molekula szintézisét eredményezi. Átlagosan 3-4 proton áthaladása szükséges egy ATP molekula szintéziséhez.
Az ATP szintáz egy lenyűgöző molekuláris gép, amely a protonok visszaáramlásának kinetikus energiáját alakítja át kémiai energiává, azaz ATP-vé, egy forgó mechanizmus segítségével.
Ezt a mechanizmust kémiozmotikus elméletnek nevezik, amelyet Peter Mitchell javasolt az 1960-as években, és amelyért 1978-ban Nobel-díjat kapott. Az elmélet forradalmasította a sejtbiológiát, megmagyarázva, hogyan kapcsolódik össze az elektron transzport és az ATP szintézis.
Az oxidatív foszforiláció szabályozása
Az oxidatív foszforiláció egy rendkívül finoman szabályozott folyamat, amelynek célja, hogy a sejt ATP-szükségletét pontosan kielégítse anélkül, hogy feleslegesen pazarolná az energiát. A szabályozás többféle szinten történik, biztosítva a rugalmasságot és az adaptációt a különböző fiziológiai állapotokhoz.
Subsztrát elérhetősége
Az oxidatív foszforiláció sebességét alapvetően befolyásolja a rendelkezésre álló szubsztrátok mennyisége. Ezek a szubsztrátok a következők:
- NADH és FADH₂: Ezek a redukált koenzimek az elektron transzport lánc „üzemanyagát” biztosítják. Mennyiségük a glikolízis, a piruvát oxidáció és a citrátkör aktivitásától függ. Ha kevés a NADH és FADH₂, az ETL lelassul.
- ADP és Pi: Az ATP szintáz működéséhez ADP és anorganikus foszfát szükséges. Ha a sejt sok energiát használ fel, az ATP hidrolizálódik ADP-re és Pi-re, ami növeli ezek koncentrációját, és serkenti az ATP szintáz működését. Ezt a jelenséget respirációs kontrollnak nevezik: az ATP/ADP arány alapvető szabályozója az oxidatív foszforilációnak. Magas ATP/ADP arány gátolja, alacsony arány serkenti a folyamatot.
- O₂: Az oxigén a végső elektronakceptor. Hiánya esetén az egész folyamat leáll.
Termék gátlás
A magas ATP-koncentráció gátolja az oxidatív foszforilációt. Ha elegendő ATP van jelen, az ATP szintáz aktivitása csökken, ami a proton gradiens növekedéséhez vezet. Ez a megnövekedett proton-mozgató erő visszahatólag lassítja az elektron transzport lánc működését is, mivel a protonok pumpálása egyre nehezebbé válik a már meglévő nagy gradiens ellenében.
Allosztérikus szabályozás
Az ETL egyes komplexei és a citrátkör enzimei is allosztérikusan szabályozottak, azaz bizonyos molekulák kötődése megváltoztatja az enzim aktivitását. Például, a citrátkör egyes lépéseit az ATP gátolja, míg az ADP serkenti, ezzel összehangolva a citrátkör és az oxidatív foszforiláció működését.
Hormonális szabályozás
Bizonyos hormonok, mint például a pajzsmirigyhormonok, befolyásolhatják a mitokondriumok számát és aktivitását, ezáltal szabályozva a sejtek anyagcseréjének általános sebességét és az oxidatív foszforiláció kapacitását. Az inzulin és a glukagon is közvetetten hat az ATP termelésre azáltal, hogy befolyásolják a glükóz és zsírsavak anyagcseréjét.
Szétkapcsoló fehérjék (Uncoupling Proteins – UCPs)
A szétkapcsoló fehérjék (UCP-k) a belső mitokondriális membránban található csatornafehérjék, amelyek lehetővé teszik a protonok visszaáramlását a mátrixba anélkül, hogy az ATP szintázon keresztül haladnának át. Ezáltal a proton gradiens energiája nem ATP szintézisére, hanem hőtermelésre fordítódik. Ez a mechanizmus különösen fontos a barna zsírszövetben, ahol az UCP1 (termogenin) szerepet játszik a testhőmérséklet fenntartásában, különösen hidegben vagy újszülötteknél. Bár az ATP termelést csökkenti, a hőtermelés létfontosságú lehet bizonyos körülmények között.
A szabályozás ezen sokrétű rendszere biztosítja, hogy a sejt energiatermelése mindig a pillanatnyi igényekhez igazodjon, elkerülve az energiafelesleget vagy a hiányt, ami károsíthatná a sejt működését.
Inhibitorok és szétkapcsolók: az oxidatív foszforiláció gátlása
Az oxidatív foszforiláció komplex folyamatának egyes lépései specifikus molekulákkal gátolhatók, ami mind a kutatásban, mind a toxikológiában, mind pedig a terápiában jelentőséggel bír. Két fő kategóriát különböztetünk meg: az inhibitorokat és a szétkapcsolókat.
Az elektron transzport lánc inhibitorai
Az inhibitorok közvetlenül az elektron transzport lánc valamelyik komplexét gátolják, megakadályozva az elektronok továbbítását és így a protonok pumpálását. Ez a proton gradiens összeomlásához és az ATP szintézis leállásához vezet.
- I. komplex inhibitorai:
- Rotenon: Növényi eredetű méreg, amelyet rovarirtóként használnak. Gátolja az elektronok továbbítását az I. komplexben a vas-kén centrumokhoz.
- Amital (barbiturát): Gyógyszer, amely szintén gátolja az I. komplexet.
Ezek az anyagok blokkolják a NADH által szállított elektronok belépését az ETL-be, de a FADH₂-ből származó elektronok (amelyek a II. komplexen keresztül lépnek be) továbbra is áramolhatnak.
- II. komplex inhibitorai:
- Malonát: Strukturálisan hasonló a szukcináthoz, kompetitív gátlója a szukcinát-dehidrogenáznak (II. komplex).
Ezek a gátlók a FADH₂ által szállított elektronok belépését akadályozzák, de a NADH-elektronok áramlása zavartalan marad.
- III. komplex inhibitorai:
- Antimicin A: Antibiotikum és gombaölő, amely a citokróm bc₁ komplexben (III. komplex) gátolja az elektronok továbbítását.
- IV. komplex inhibitorai:
- Cianid (CN⁻): Erős méreg, amely irreverzibilisen kötődik a citokróm c oxidáz (IV. komplex) vasatomjához, megakadályozva az oxigénhez való elektronátadást. Ez a legkritikusabb gátlás, mivel az egész lánc leáll, és az ATP termelés szinte teljesen megszűnik.
- Szén-monoxid (CO): Hasonlóan a cianidhoz, a IV. komplex hemjéhez kötődik, kompetitíven gátolva az oxigén kötődését.
- Azid (N₃⁻): Szintén a IV. komplexet gátolja.
Ezek az anyagok rendkívül toxikusak, mivel az aerob légzés kulcsfontosságú lépését blokkolják, ami gyors sejthalálhoz vezet.
- ATP szintáz inhibitorai:
- Oligomicin: Ez az antibiotikum az ATP szintáz F₀ alegységéhez kötődik, és blokkolja a protonok áramlását a csatornán keresztül. Ennek következtében a proton gradiens megnő, de az ATP szintézis nem tud végbemenni. A magas proton gradiens visszahat az ETL-re, és lassítja az elektronok áramlását is.
Szétkapcsolók (Uncouplers)
A szétkapcsolók olyan vegyületek, amelyek a proton gradienst megszüntetik anélkül, hogy közvetlenül gátolnák az elektron transzport láncot. Ezek a molekulák hidrofóbak és képesek protonokat szállítani a belső mitokondriális membránon keresztül, bypassingolva az ATP szintázt. Ennek eredményeként az elektron transzport folytatódik, a protonok pumpálása is megtörténik, de a protonok azonnal visszaszivárognak a mátrixba, így a gradiens nem tud felépülni. Az energia hő formájában disszipálódik, és az ATP szintézis drasztikusan csökken vagy leáll.
- 2,4-dinitrofenol (DNP): Ez a klasszikus szétkapcsoló egy lipidoldékony molekula, amely képes protonokat felvenni a membránok közötti térben, áthaladni a belső membránon, majd leadni azokat a mátrixban. Korábban súlycsökkentő szerként használták, de rendkívül veszélyes mellékhatásai (kontrollálatlan hőtermelés, hipertermia, halál) miatt betiltották.
- Valinomicin: Antibiotikum, amely káliumionokat szállít a membránon keresztül, és közvetetten befolyásolja a membránpotenciált.
- Gramicidin: Egy másik antibiotikum, amely ioncsatornákat képez a membránban, lehetővé téve a protonok és más ionok áramlását.
Az inhibitorok és szétkapcsolók tanulmányozása alapvetően hozzájárult az oxidatív foszforiláció mechanizmusának megértéséhez. Emellett potenciális terápiás célpontokat is jelentenek bizonyos betegségek, például metabolikus rendellenességek vagy rák kezelésében, bár alkalmazásuk komoly mellékhatásokkal járhat.
Az oxidatív foszforiláció élettani jelentősége
Az oxidatív foszforiláció az aerob szervezetek, így az emberi test számára is az elsődleges ATP-forrás. Élettani jelentősége rendkívül sokrétű, és az egész szervezet működésére kihat.
Az ATP termelés hatékonysága
Az oxidatív foszforiláció kiemelkedő hatékonyságával messze felülmúlja a szubsztrát-szintű foszforilációt. Míg egy glükózmolekula anaerob lebontásával (glikolízis) mindössze 2 ATP keletkezik, addig az aerob légzés (glikolízis, piruvát oxidáció, citrátkör és oxidatív foszforiláció) során akár 30-32 ATP is termelődhet. Ez a nagyfokú energiahozam teszi lehetővé a magas energiaigényű folyamatok fenntartását a szervezetben.
Szerepe a különböző szövetekben
Minden sejtnek szüksége van ATP-re, de egyes szövetek energiaigénye különösen magas, így az oxidatív foszforiláció ott kiemelt szerepet kap:
- Izomszövet: A vázizomzat, különösen a lassan rángatózó (oxidatív) rostok, nagymértékben támaszkodnak az oxidatív foszforilációra a folyamatos mozgáshoz és a hosszan tartó terheléshez szükséges ATP előállításában. A szívizom szinte kizárólag aerob módon termeli az energiát.
- Agy: Az agy rendkívül energiaigényes szerv, a szervezet oxigénfogyasztásának mintegy 20%-át teszi ki. Az idegsejtek folyamatosan ATP-t használnak fel az ionpumpák működtetéséhez, a neurotranszmitterek szintéziséhez és a szinaptikus átvitelhez. Az agyban az oxidatív foszforiláció zavara súlyos neurológiai problémákhoz vezethet.
- Máj: A máj számos anyagcsere-folyamat központja, beleértve a glükóz szintézisét (glükoneogenezis), a zsírsav-anyagcserét és a méregtelenítést, amelyek mind jelentős ATP-felhasználással járnak.
- Vesék: A vesék folyamatosan szűrik a vért és visszaszívják a hasznos anyagokat, ami energiaigényes transzportfolyamatokat igényel.
Hőtermelés (termogenezis)
Ahogy korábban említettük, a szétkapcsoló fehérjék (UCP-k) lehetővé teszik a proton gradiens energiájának hővé alakítását. Ez a mechanizmus különösen fontos a testhőmérséklet szabályozásában, különösen csecsemőknél, ahol a barna zsírszövetben zajló hőtermelés létfontosságú a hideg elleni védekezésben. Felnőtteknél is szerepet játszik a testhőmérséklet fenntartásában és az energiaegyensúly szabályozásában.
Reaktív oxigénfajták (ROS) termelése
Bár az oxidatív foszforiláció rendkívül hatékony, nem tökéletes. Az elektron transzport lánc működése során az oxigén nem mindig redukálódik teljesen vízzé. Időnként, főleg a I. és III. komplexeknél, egy elektron „elszökhet” és részlegesen redukálhatja az oxigénmolekulát, szuperoxid gyököt (O₂⁻) képezve. Ez az első reaktív oxigénfajta (ROS), amely további, rendkívül káros ROS-okká alakulhat (pl. hidrogén-peroxid, hidroxil gyök).
- ROS termelés mechanizmusa: A részlegesen redukált oxigénmolekulák rendkívül reaktívak, és károsíthatják a sejtek makromolekuláit, mint például a DNS-t, fehérjéket és lipideket. Ez az oxidatív stressz számos betegség és az öregedési folyamat alapja.
- Antioxidáns védekezés: A sejtek számos antioxidáns enzimmel (pl. szuperoxid-diszmutáz, kataláz, glutation-peroxidáz) és nem enzimatikus antioxidánssal (pl. C-vitamin, E-vitamin, glutation) rendelkeznek, amelyek semlegesítik a ROS-okat és védelmet nyújtanak az oxidatív károsodás ellen.
Az oxidatív foszforiláció tehát nem csak az energiaellátás motorja, hanem egy potenciális forrása is a sejtek károsodásának, ami rávilágít a precíz szabályozás és a kiegyensúlyozott működés fontosságára.
Oxidatív foszforiláció és betegségek

Az oxidatív foszforiláció központi szerepe a sejtek energiaellátásában azt jelenti, hogy bármilyen zavar a folyamatban súlyos következményekkel járhat az egészségre. Számos betegség közvetlenül vagy közvetetten kapcsolódik a mitokondriális diszfunkcióhoz és az oxidatív foszforiláció zavaraihoz.
Mitokondriális betegségek
Ezek a betegségek a mitokondriumok genetikai vagy szerzett hibáiból erednek, amelyek befolyásolják az oxidatív foszforiláció komponenseinek működését. Mivel a mitokondriumoknak saját DNS-e is van, a hibák származhatnak mind a nukleáris, mind a mitokondriális genomból. Jellemzően azokat a szöveteket érintik legsúlyosabban, amelyeknek magas az energiaigénye, mint például az agy, az izmok, a szív és a máj.
- Leigh-szindróma: Progresszív neurodegeneratív rendellenesség, amelyet az oxidatív foszforiláció komplexek, különösen a I. és IV. komplex, zavarai okoznak.
- MELAS-szindróma (Mitochondrial Encephalomyopathy, Lactic Acidosis, and Stroke-like episodes): Jellemzően a mitokondriális DNS mutációi okozzák, amelyek a légzési lánc fehérjéinek szintézisét befolyásolják.
- Kearns-Sayre-szindróma: Nagy mitokondriális DNS deléciók jellemzik, amelyek számos mitokondriális gén működését érintik.
A tünetek széles skálán mozoghatnak, beleértve az izomgyengeséget, neurológiai problémákat, szívbetegségeket, veseelégtelenséget és endokrin rendellenességeket. A kezelés gyakran tüneti, és a mitokondriális funkciót támogató kiegészítőket (pl. koenzim Q10) alkalmaznak.
Neurodegeneratív betegségek
Az oxidatív foszforiláció zavarai kulcsszerepet játszhatnak az olyan neurodegeneratív betegségek kialakulásában, mint a Parkinson-kór és az Alzheimer-kór.
- Parkinson-kór: A dopaminerg neuronok pusztulásával jár a substantia nigra területén. Számos tanulmány kimutatta a I. komplex aktivitásának csökkenését a Parkinson-kóros betegek agyában, ami megnövekedett oxidatív stresszhez és sejthalálhoz vezethet.
- Alzheimer-kór: Jellemzője az amiloid plakkok és tau-fehérje gomolygok felhalmozódása az agyban. Az oxidatív foszforiláció zavarai, a mitokondriális diszfunkció és az oxidatív stressz jelentősen hozzájárulnak a neuronok károsodásához és a betegség progressziójához.
Rák és metabolikus szindróma
A rákos sejtek metabolizmusa gyakran eltér a normál sejtekétől, amit Warburg-effektusnak neveznek. A rákos sejtek még oxigén jelenlétében is inkább anaerob glikolízist végeznek, mint oxidatív foszforilációt, bár az utóbbi sokkal hatékonyabb ATP-termelő. Ez a metabolikus átprogramozás lehetővé teszi a rákos sejtek gyors proliferációját és a biomassza felépítését. Az oxidatív foszforiláció gátlása vagy modulálása potenciális terápiás stratégiát jelenthet a rák kezelésében.
A metabolikus szindróma, amely magában foglalja az elhízást, inzulinrezisztenciát, magas vérnyomást és diszlipidémiát, szintén kapcsolódik a mitokondriális diszfunkcióhoz. Az elhízott és inzulinrezisztens egyénekben gyakran megfigyelhető az oxidatív foszforiláció hatékonyságának csökkenése, ami hozzájárulhat az energiaegyensúly felborulásához és az oxidatív stresszhez.
Öregedés
Az öregedési folyamat egyik vezető elmélete a mitokondriális károsodás felhalmozódása. Az oxidatív foszforiláció során termelődő ROS-ok hosszú távon károsíthatják a mitokondriális DNS-t, fehérjéket és lipideket, ami a mitokondriumok működésének romlásához vezet. Ez az energiaellátás csökkenését, a ROS-termelés további növekedését és a sejtek funkciójának hanyatlását eredményezi, hozzájárulva az öregedés látható jeleihez és az életkorral összefüggő betegségekhez.
A mitokondriális diszfunkció és az oxidatív foszforiláció zavarai tehát számos, a modern társadalmat sújtó betegség patogenezisében játszanak szerepet, hangsúlyozva a folyamat alapos megértésének és a terápiás beavatkozások kidolgozásának fontosságát.
Összehasonlítás a szubsztrát-szintű foszforilációval
Bár mindkét folyamat ATP-t termel, az oxidatív foszforiláció és a szubsztrát-szintű foszforiláció alapvetően eltérő mechanizmusokon keresztül működik, és különböző szerepeket tölt be a sejt anyagcseréjében.
Szubsztrát-szintű foszforiláció
- Mechanizmus: Közvetlenül, egy enzim segítségével egy magas energiájú foszfátcsoportot tartalmazó szubsztrát molekuláról visz át egy foszfátcsoportot az ADP-re, ATP-t képezve. Nincs szükség elektron transzport láncra vagy proton gradiensre.
- Helyszín: Elsősorban a citoplazmában (glikolízis során) és a mitokondrium mátrixában (citrátkör során).
- Oxigénfüggőség: Nem igényel oxigént. Anaerob körülmények között is képes ATP-t termelni.
- ATP hozam: Viszonylag alacsony. A glikolízis során 2 ATP keletkezik, a citrátkörben pedig 2 GTP (ami ATP-vé alakítható).
- Sebesség: Gyorsabb, azonnali ATP-ellátást biztosít a sejtnek, de korlátozott mennyiségben.
- Fiziológiai szerep: Fontos az azonnali energiaigények fedezésében, különösen oxigénhiányos állapotokban (pl. intenzív izommunka során). Gyorsan mobilizálható energiaforrás.
Oxidatív foszforiláció
- Mechanizmus: Indirekt módon termel ATP-t. Az elektron transzport láncban az elektronok áramlása során felszabaduló energiát proton gradiens létrehozására használja fel, amelyet az ATP szintáz hasznosít az ATP szintézisére.
- Helyszín: Mitokondrium belső membránja.
- Oxigénfüggőség: Szigorúan oxigénfüggő, mivel az oxigén a végső elektronakceptor.
- ATP hozam: Rendkívül magas. Egy glükózmolekula teljes oxidációjából akár 30-32 ATP is keletkezhet.
- Sebesség: Lassabb, mint a szubsztrát-szintű foszforiláció, de folyamatos és nagy mennyiségű ATP-t biztosít.
- Fiziológiai szerep: Az aerob szervezetek elsődleges ATP-forrása, fedezi a sejtek hosszú távú, nagy energiaigényét, és fenntartja az alapvető életfolyamatokat.
A két folyamat tehát kiegészíti egymást. A szubsztrát-szintű foszforiláció gyors, de korlátozott energiát biztosít, míg az oxidatív foszforiláció lassabb, de rendkívül hatékony és folyamatos energiaellátást garantál az aerob körülmények között működő sejtek számára. Az élet fenntartásához mindkét mechanizmus elengedhetetlen, és a sejt képes adaptálni az arányukat a pillanatnyi energiaigény és oxigénellátottság függvényében.
Jövőbeli kutatási irányok és terápiás potenciál
Az oxidatív foszforiláció és a mitokondriális funkciók mélyebb megértése folyamatosan új utakat nyit meg a gyógyászatban és a biológiai kutatásban. A folyamat komplexitása és központi szerepe miatt számos betegség kezelésében rejt potenciált.
Mitokondriális gyógyszerek fejlesztése
A mitokondriális betegségek, neurodegeneratív kórképek és a metabolikus szindróma kezelésére irányuló kutatások egyre inkább a mitokondriumokra és az oxidatív foszforilációra fókuszálnak. A gyógyszerfejlesztés célja lehet:
- Az elektron transzport lánc komplexek aktivitásának modulálása: Olyan vegyületek keresése, amelyek serkentik az alulműködő komplexek aktivitását, vagy gátolják a túlműködő, ROS-termelő komplexeket.
- ATP szintáz funkciójának optimalizálása: Az ATP termelés hatékonyságának növelése vagy a diszfunkcionális ATP szintázok korrigálása.
- Antioxidáns védekezés erősítése: Célzott antioxidánsok, amelyek képesek bejutni a mitokondriumokba és ott semlegesíteni a ROS-okat, csökkentve az oxidatív stresszt. Példaként említhető a MitoQ, egy mitokondrium-célzott koenzim Q10 analóg.
- Mitokondriális biogenezis serkentése: Olyan molekulák, amelyek növelik a mitokondriumok számát vagy javítják minőségüket a sejtekben, ezzel növelve az oxidatív foszforiláció kapacitását.
Génterápia és mitokondriális transzplantáció
A mitokondriális betegségek genetikai eredete miatt a génterápia ígéretes megközelítés lehet. Célja a hibás gének kijavítása vagy funkcionális kópiák bejuttatása a sejtbe. A mitokondriális transzplantáció, ahol egészséges mitokondriumokat juttatnak be a beteg sejtekbe, szintén intenzív kutatás tárgya, különösen olyan állapotokban, mint a szívroham vagy az agyvérzés.
Az oxidatív foszforiláció és az immunválasz
Egyre több bizonyíték utal arra, hogy a mitokondriális metabolizmus, beleértve az oxidatív foszforilációt is, kulcsszerepet játszik az immunsejtek működésének szabályozásában. Különböző immunsejtek (pl. T-sejtek, makrofágok) különböző metabolikus programokat használnak a funkciójuktól függően. Ennek a kapcsolatnak a megértése új terápiás lehetőségeket nyithat meg autoimmun betegségek vagy rákos megbetegedések kezelésében.
Az öregedés elleni küzdelem
Az oxidatív foszforiláció, a ROS termelés és a mitokondriális diszfunkció közötti kapcsolat miatt a mitokondriumok az öregedés elleni stratégiák egyik fő célpontjai. A mitokondriális funkciók optimalizálása, az oxidatív stressz csökkentése és a mitofágia (a sérült mitokondriumok eltávolítása) serkentése kulcsfontosságú lehet az egészséges öregedés elősegítésében és az életkorral összefüggő betegségek megelőzésében.
Az oxidatív foszforiláció tehát nem csupán egy alapvető biokémiai folyamat, hanem egy dinamikus rendszer, amelynek zavarai széles spektrumú betegségekhez vezethetnek. A folyamatos kutatás reményt ad arra, hogy a jövőben hatékonyabb diagnosztikai és terápiás eszközök állnak majd rendelkezésre ezen komplex mechanizmusok kezelésére.







