
Az élővilág egyik legősibb és legfundamentálisabb jelensége a sejtek belső környezetének precíz szabályozása. Ahhoz, hogy egy sejt életben maradjon, működjön és kommunikáljon a külvilággal, folyamatosan fenn kell tartania a belső és külső ionkoncentrációk közötti különbségeket. Ezen komplex és dinamikus egyensúly fenntartásának egyik legfontosabb, ha nem a legfontosabb motorja egy molekuláris csoda: a Na+/K+-ATPáz. Ez az enzim nem csupán egy egyszerű pumpa, hanem egy elektrogén transzporter, amely a sejtek energiahordozóját, az adenozin-trifoszfátot (ATP) felhasználva képes a nátrium- és káliumionokat aktívan mozgatni a sejtmembránon keresztül, a koncentrációgrádienssel szemben. Ezzel alapvetően hozzájárul a nyugalmi membránpotenciál kialakításához, a sejtvolumen szabályozásához, és közvetve számos más, létfontosságú élettani folyamathoz, a tápanyagfelvételtől az idegi ingerületvezetésig.
Az Na+/K+-ATPáz felfedezése és története
Az Na+/K+-ATPáz felfedezése egy dán tudós, Jens Christian Skou nevéhez fűződik, aki az 1950-es években a nátrium és kálium ionok aktív transzportjának mechanizmusát vizsgálta rákidegsejtekben. Skou professzor felismerte, hogy a nátrium- és káliumionok mozgása a sejtmembránon keresztül ATP-függő, és az ATP hidrolíziséért felelős enzim aktivitása szorosan összefügg ezen ionok jelenlétével. Kísérletei során kimutatta, hogy az ATP-hidrolízis csak akkor optimális, ha mind nátrium-, mind káliumionok jelen vannak a megfelelő oldalon – nátrium a sejt belsejében, kálium a külsejében. Ez a forradalmi felfedezés alapozta meg a modern sejtélettan és membránbiológia egyik legfontosabb pillérét, és Skou professzornak 1997-ben orvosi-élettani Nobel-díjat hozott.
Skou munkássága rávilágított arra, hogy a sejtek nem passzív, hanem aktív módon képesek fenntartani az iongrádienseket, ami elengedhetetlen a homeosztázis és a sejtfunkciók szempontjából. Az azóta eltelt évtizedekben az Na+/K+-ATPáz szerkezetét, működését és élettani szerepét részletesen feltárták, bebizonyítva, hogy ez az enzim az élet egyik alapköve. A kezdeti biokémiai és élettani vizsgálatoktól a modern molekuláris biológiai és szerkezeti elemzésekig, az enzimről szerzett ismereteink folyamatosan bővültek, és ma már tudjuk, hogy az emberi test minden egyes sejtjében jelen van, és nélkülözhetetlen a túléléshez.
„A nátrium-kálium pumpa a sejtek energiahordozóját, az ATP-t felhasználva tartja fenn az iongrádienseket, melyek alapvetőek az élethez.”
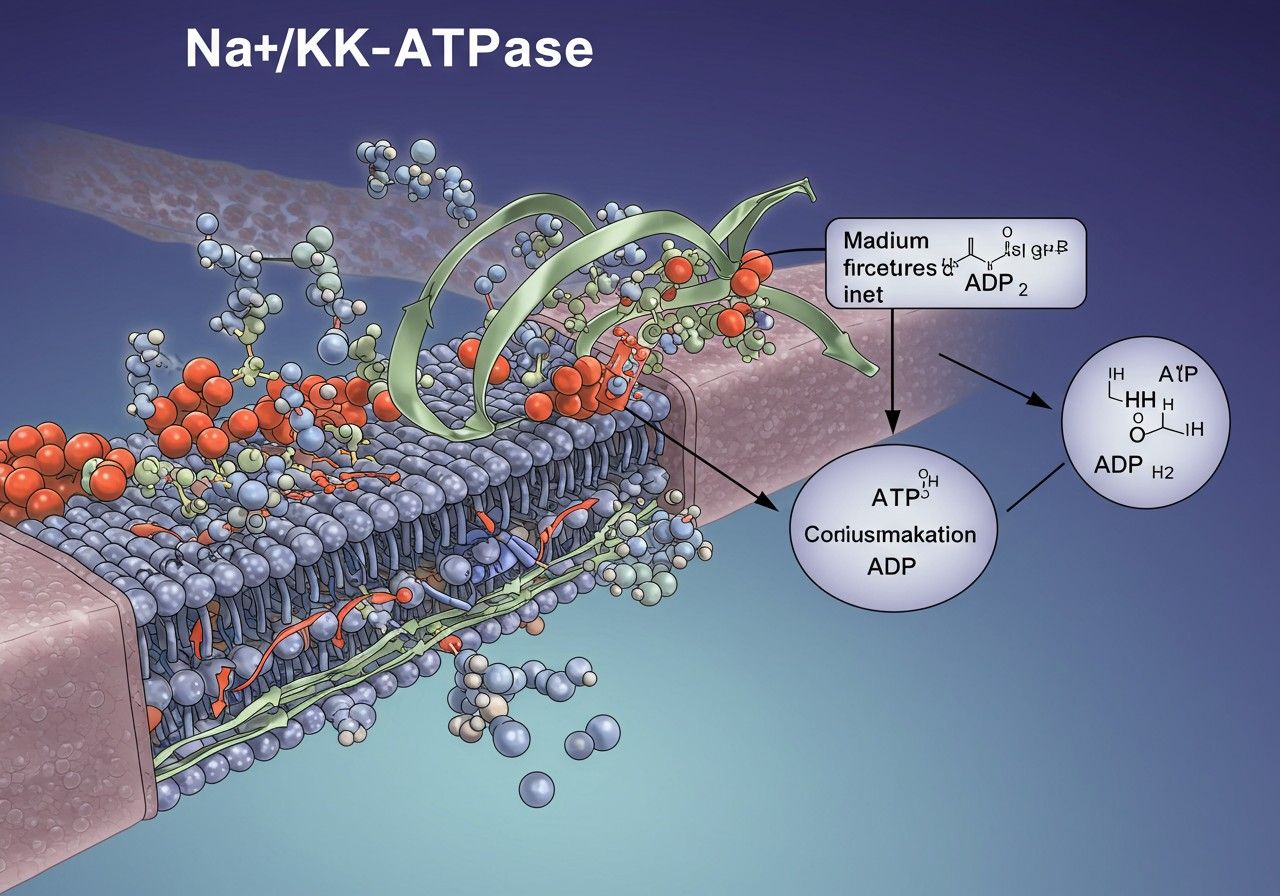
Az enzim szerkezete és felépítése
Az Na+/K+-ATPáz egy komplex transzmembrán fehérje, amely több alegységből épül fel, ezek közül a legfontosabbak az alfa (α) és a béta (β) alegységek, valamint bizonyos izoformákban egy gamma (γ) alegység is. Ezek az alegységek szinergikusan működnek együtt, hogy biztosítsák az enzim hatékony és specifikus működését.
Az alfa (α) alegység a pumpa „motorja” és a funkcionalitás magja. Ez egy nagy méretű (kb. 1000 aminosavból álló) fehérje, amely számos transzmembrán doménnel rendelkezik, átszelve a sejtmembránt. Az α alegység tartalmazza az összes kulcsfontosságú funkcionális helyet: a nátrium- és káliumkötő helyeket, az ATP-kötő helyet, valamint a foszforilációs helyet. Ez az alegység felelős az ATP hidrolíziséért és a konformációs változásokért, amelyek az ionok transzportját eredményezik. Az emberi szervezetben több izoformája (α1, α2, α3, α4) ismert, melyek eltérő szöveti eloszlással és kinetikai tulajdonságokkal rendelkeznek, lehetővé téve a specifikus élettani szerepeket. Például az α1 izoformát szinte minden sejtben megtaláljuk, míg az α2 az izmokban és a szívben, az α3 az idegrendszerben, az α4 pedig a hímivarsejtekben domináns.
A béta (β) alegység egy glikozilezett transzmembrán fehérje, amelynek funkciója elsősorban az α alegység membránba való beépülésének, stabilitásának és megfelelő működésének biztosítása. Noha önmagában nincs ATP-hidrolizáló vagy iontranszportáló képessége, elengedhetetlen az α alegység helyes hajtogatásához és szállításához a sejtmembránhoz. A β alegységnek szintén több izoformája (β1, β2, β3) ismert, melyek az α alegységekkel hasonlóan szövet-specifikus expressziót mutatnak. A glikozilációja szerepet játszhat a sejt-sejt interakciókban és a sejtfelszíni felismerési folyamatokban is.
A gamma (γ) alegység egy kisebb, egyetlen transzmembrán doménnel rendelkező fehérje, amelyet csak bizonyos izoformákhoz társulva találtak meg, például a vesében és a szívben. A γ alegység nem minden Na+/K+-ATPáz komplex része, és szerepe a pumpa aktivitásának modulálásában, finomhangolásában rejlik. Képes befolyásolni a pumpa affinitását a nátrium- és káliumionokhoz, valamint az ATP-hez, ezzel módosítva a transzport sebességét és hatékonyságát. Ezen alegységek komplex együttműködése biztosítja, hogy a Na+/K+-ATPáz a sejtek ionhomeosztázisának sarokköveként funkcionálhasson, alkalmazkodva a különböző szövetek és élettani állapotok specifikus igényeihez.
A működési mechanizmus részletesen
Az Na+/K+-ATPáz működése egy ciklikus folyamat, amelyet a konformációs változások jellemeznek, és két fő állapot, az E1 és E2 konformáció közötti váltakozáson alapul. Ez a ciklus biztosítja a nátriumionok kiáramlását és a káliumionok beáramlását a sejtbe, az ATP energiájának felhasználásával.
A ciklus az E1 konformációval kezdődik, amikor az enzim nagy affinitással rendelkezik a nátriumionok iránt a sejt belsejében. Ebben az állapotban három nátriumion (3 Na+) kötődik az enzim intracelluláris oldalán lévő kötőhelyekhez. A nátriumionok kötődése elősegíti az ATP kötődését az enzimhez.
Az ATP kötődése után az enzim saját magát foszforilálja egy aszparaginsav oldalláncon, az ATP terminális foszfátcsoportjának átvitelével. Ez a foszforiláció egy energialökés, amely a pumpa szerkezetében jelentős változást, az E1-ből E2 konformációba való átalakulást váltja ki. Az E2 konformációban a nátriumkötő helyek affinitása drasztikusan lecsökken, és a kötőhelyek kifelé, a sejten kívüli tér felé nyílnak. Ennek következtében a három nátriumion felszabadul a sejt külső oldalán.
Miután a nátriumionok távoztak, az E2 konformációban lévő enzim nagy affinitással rendelkezik a káliumionok iránt a sejt külső oldalán. Ekkor két káliumion (2 K+) kötődik az enzim extracelluláris oldalán lévő kötőhelyekhez. A káliumionok kötődése kiváltja a foszfátcsoport hidrolitikus lehasadását az enzimről (de-foszforiláció). Ez a de-foszforiláció egy újabb konformációs változást idéz elő, visszaállítva az enzimet az E1 állapotba.
Az E1 konformációban a káliumkötő helyek affinitása lecsökken, és a kötőhelyek befelé, a sejt belseje felé nyílnak. Ennek eredményeként a két káliumion felszabadul az intracelluláris térben. Ezzel a ciklus bezárul, és az enzim készen áll egy újabb körre. Fontos kiemelni, hogy minden egyes ciklus során 3 Na+ ion távozik a sejtből és 2 K+ ion jut be, ami nettó pozitív töltés kiáramlását jelenti. Ezért nevezzük az Na+/K+-ATPázt elektrogén pumpának, mivel közvetlenül hozzájárul a sejtmembránon átívelő elektromos potenciálkülönbség, a nyugalmi membránpotenciál fenntartásához. Az ATP hidrolíziséből származó energia biztosítja, hogy ez a folyamat a koncentráció- és elektromos grádienssel szemben is folyamatosan működhessen, fenntartva a sejtek számára létfontosságú iongrádienseket.
Energetikai szempontok: miért ATP?

Az Na+/K+-ATPáz működésének alapja az adenozin-trifoszfát (ATP) hidrolíziséből felszabaduló kémiai energia hasznosítása. Az ATP nem véletlenül a sejtek univerzális energiavalutája; nagy energiájú foszfátkötéseinek hidrolízise jelentős mennyiségű energiát szabadít fel, amely alkalmassá teszi a sejten belüli energiaigényes folyamatok hajtására.
A nátrium- és káliumionok transzportja a sejtmembránon keresztül egyértelműen aktív transzport, mivel az ionok a koncentrációgrádienssel szemben mozognak. A nátriumionok intracelluláris koncentrációja alacsonyabb, mint az extracelluláris koncentrációja, míg a káliumionok esetében éppen fordítva van: magasabb az intracelluláris, mint az extracelluláris koncentráció. Ezeknek a grádienseknek a fenntartása folyamatos energia ráfordítást igényel, mivel az ionok passzívan, ioncsatornákon keresztül igyekeznének kiegyenlítődni.
Az ATP hidrolízise során a terminális foszfátcsoport lehasadása (ATP → ADP + Pi) körülbelül -30.5 kJ/mol standard szabadenergia-változással jár. Ez a felszabaduló energia elegendő ahhoz, hogy a pumpa konformációs változásait meghajtsa, és az ionokat a grádienssel szemben mozgassa. Az enzim foszforilációja, majd de-foszforilációja során ez az energia átmenetileg az enzim szerkezetébe tárolódik, majd felszabadul, lehetővé téve a kötőhelyek affinitásának és orientációjának változását. Az Na+/K+-ATPáz rendkívül hatékonyan alakítja át a kémiai energiát mechanikai munkává, biztosítva a sejtek ionhomeosztázisának fenntartását még extrém körülmények között is. A sejt ATP-termelésének jelentős része, becslések szerint akár 20-40%-a is erre az egyetlen enzimre fordítódik, ami jól mutatja annak létfontosságú szerepét az energiafelhasználásban és a sejtélettanban.
Élettani szerepe a sejtekben
Az Na+/K+-ATPáz élettani szerepe annyira alapvető és szerteágazó, hogy gyakorlatilag minden sejt és szövet működéséhez elengedhetetlen. Nélküle a sejtek képtelenek lennének fenntartani integritásukat és funkcióikat.
Membránpotenciál fenntartása
Az egyik legközvetlenebb és legfontosabb szerepe a nyugalmi membránpotenciál kialakításában és fenntartásában rejlik. Mivel az enzim minden ciklusban 3 Na+ iont pumpál ki a sejtből és 2 K+ iont pumpál be, nettó egy pozitív töltés távozik a sejtből. Ez az elektrogén hatás közvetlenül hozzájárul a sejt belsejének negatívabb töltéséhez, ami a nyugalmi membránpotenciál alapját képezi. Bár az ioncsatornákon keresztül történő passzív ionáramlások is jelentősen befolyásolják a potenciált, a pumpa folyamatos munkája biztosítja, hogy a grádiensek ne egyenlítődjenek ki, és a sejt mindig készen álljon az ingerületvezetésre, például az ideg- és izomsejtek esetében.
Sejtvolumen szabályozása
Az Na+/K+-ATPáz létfontosságú szerepet játszik a sejtvolumen szabályozásában. A sejt belsejében lévő magasabb fehérje- és más szerves anionkoncentráció miatt a sejt ozmotikusan hajlamos lenne vizet felvenni és megduzzadni. A pumpa folyamatosan eltávolítja a nátriumot a sejtből, csökkentve ezzel az intracelluláris ozmotikusan aktív részecskék számát. Ez a nátriumgradiens hozzájárul ahhoz, hogy a víz passzívan kövesse az ionokat, és kifelé áramoljon a sejtből, megakadályozva a sejt duzzadását és lízisét. Egy működésképtelen Na+/K+-ATPáz esetén a sejtek gyorsan megduzzadnának és elpusztulnának.
Másodlagos aktív transzport hajtóereje
Az Na+/K+-ATPáz által fenntartott nátriumgradiens egy hatalmas energiatartalékot képvisel, amelyet a sejtek számos másodlagos aktív transzportfolyamat hajtására használnak fel. A másodlagos aktív transzport során egy másik molekula (pl. glükóz, aminosav) a nátriummal együtt, a nátrium grádiensének irányába mozog be a sejtbe (kotranszport), vagy a nátriummal ellentétes irányba távozik a sejtből (kontratranszport), miközben a nátrium befelé áramlik.
Jellemző példa erre a glükóz és aminosavak felvétele a bélhámsejtekben és a vesetubulusokban (SGLT transzporterek). Ezek a transzporterek a nátrium befelé áramlásának energiáját használják fel a glükóz vagy aminosavak koncentrációgrádienssel szembeni felvételéhez. Hasonlóképpen, a Na+/Ca2+ kicserélő (NCX) a szívizomsejtekben a nátrium befelé áramlását használja fel a kalcium kiáramoltatására, ami létfontosságú a szívizom relaxációjához és a kontrakciós ciklus szabályozásához. Az Na+/K+-ATPáz tehát közvetve biztosítja a sejtek tápanyagellátását és a kalcium homeosztázisát.
Idegsejt működés
Az idegsejtek működésében az Na+/K+-ATPáz alapvető szerepet játszik az akciós potenciál kialakulása és terjedése utáni repolarizációban és a nyugalmi membránpotenciál helyreállításában. Az akciós potenciál során nagyszámú Na+ ion áramlik be a sejtbe, és K+ ionok áramlanak ki. Az Na+/K+-ATPáz feladata, hogy ezeket az ionokat visszapumpálja eredeti helyükre, ezzel helyreállítva az iongrádienseket és biztosítva az idegsejt újabb ingerelhetőségét. Nélküle az idegsejtek képtelenek lennének sorozatosan akciós potenciálokat generálni, ami az idegrendszer működésének leállásához vezetne.
Izomkontrakció
Az izomsejtekben, különösen a szívizomban, az Na+/K+-ATPáz kulcsfontosságú a kontrakció és relaxáció megfelelő üteméhez. Fenntartja a nátriumgrádienst, ami lehetővé teszi a Na+/Ca2+ kicserélő (NCX) működését. Az NCX a nátrium befelé áramlásával kalciumot pumpál ki a sejtből, csökkentve az intracelluláris kalciumszintet, ami elengedhetetlen az izom relaxációjához. Ha az Na+/K+-ATPáz aktivitása csökken, az intracelluláris nátrium szintje megemelkedik, ami gátolja az NCX működését, kevesebb kalcium ürül ki a sejtből, és ez fokozottabb, hosszan tartó kontrakciót eredményezhet. Ez az elv áll a szívglikozidok hatásmechanizmusának hátterében is.
Vese működése
A vesékben az Na+/K+-ATPáz rendkívül magas koncentrációban van jelen, különösen a tubuláris sejtek basolaterális membránján. Itt alapvető szerepet játszik a nátrium, kálium, glükóz, aminosavak és víz reabszorpciójában. Az Na+/K+-ATPáz folyamatosan pumpálja a nátriumot a tubuláris sejtekből az intersticiális térbe, létrehozva egy nátriumgrádienst, amelyet aztán a szomszédos lumenális membránon található kotranszporterek (pl. SGLT, Na+/H+ kicserélő) használnak fel a szűrletből származó ionok és tápanyagok visszaszállítására a vérbe. A vese működésének mintegy 80%-a az Na+/K+-ATPáz aktivitásától függ, ami rávilágít az enzim kardinális szerepére a folyadék- és elektrolit-egyensúly fenntartásában.
Spermatozoon mozgás
A spermatozoonok mozgásképességéhez és érési folyamataihoz is nélkülözhetetlen az Na+/K+-ATPáz. A spermiumok farokrészében, a flagellum membránjában expresszálódó specifikus izoformák hozzájárulnak a membránpotenciál fenntartásához, ami kulcsfontosságú a motilitáshoz szükséges ioncsatornák és transzporterek működéséhez. Az iongrádiensek befolyásolják a spermiumok kapacitációját és az akroszóma reakciót is, amelyek elengedhetetlenek a megtermékenyítéshez.
A gyomor savtermelése
Bár a gyomor savtermeléséért közvetlenül a H+/K+-ATPáz (protonpumpa) felelős, az Na+/K+-ATPáz indirekt módon mégis részt vesz ebben a folyamatban. A parietális sejtekben az Na+/K+-ATPáz fenntartja az intracelluláris nátriumgrádienst, amely lehetővé teszi a Na+/H+ kicserélő működését. Ez a kicserélő a nátrium beáramlásával hidrogénionokat pumpál ki a sejtből, amelyek végül a H+/K+-ATPáz által kerülnek a gyomor lumenébe. Ezen túlmenően a megfelelő iongrádiensek elengedhetetlenek a sejtek általános életképességéhez és funkciójához, így a parietális sejtek is csak az Na+/K+-ATPáz működésével képesek hatékonyan savat termelni.
Az Na+/K+-ATPáz szabályozása
Az Na+/K+-ATPáz aktivitása nem állandó, hanem a sejt és a szervezet igényeihez igazodva precízen szabályozott. Számos tényező befolyásolhatja az enzim működését, biztosítva a rugalmas alkalmazkodást a változó élettani körülményekhez.
Hormonális szabályozás
Számos hormon képes modulálni az Na+/K+-ATPáz aktivitását, gyakran a génexpresszió vagy a transzporter membránba való beépülésének befolyásolásával.
- Aldoszteron: Ez a mineralokortikoid hormon a vesetubulusokban (gyűjtőcsatornákban) fokozza a nátrium reabszorpciót és a kálium szekréciót. Az aldoszteron növeli az Na+/K+-ATPáz szintézisét és membránba történő beépülését, ezzel fokozva az enzim aktivitását és a nátrium visszaszívását.
- Pajzsmirigyhormonok (T3, T4): A pajzsmirigyhormonokról ismert, hogy növelik az alapanyagcserét és a hőtermelést. Ennek egyik mechanizmusa az Na+/K+-ATPáz aktivitásának fokozása, ami több ATP fogyasztásával jár, és így hőtermeléshez vezet.
- Inzulin: Az inzulinról kimutatták, hogy serkenti az Na+/K+-ATPáz aktivitását bizonyos sejtekben, például az izom- és zsírsejtekben. Ez hozzájárul a kálium felvételéhez a sejtekbe, ami fontos a hiperkalémia megelőzésében.
- Katekolaminok (adrenalin, noradrenalin): Ezek a stresszhormonok is befolyásolhatják az enzim aktivitását, főleg a szívben és az erekben, a cAMP-függő jelátviteli útvonalakon keresztül.
Neurotranszmitterek
Az idegrendszerben a neurotranszmitterek, mint például a dopamin vagy az acetilkolin, szintén képesek modulálni az Na+/K+-ATPáz működését, befolyásolva az idegsejtek ingerlékenységét és a szinaptikus átvitelt. Ez a moduláció történhet közvetlenül, az enzimhez kötődve, vagy közvetetten, másodlagos hírvivő rendszereken keresztül.
Intracelluláris tényezők
A sejten belüli környezet számos paramétere közvetlenül befolyásolja az enzim működését:
- Intracelluláris Na+ és extracelluláris K+ koncentráció: Az enzim aktivitása szorosan függ szubsztrátjainak, azaz a nátrium- és káliumionoknak a rendelkezésre állásától. Magas intracelluláris Na+ és magas extracelluláris K+ koncentráció fokozza a pumpa aktivitását, mivel nagyobb a „meghajtóerő” az ionok transzportjára.
- ATP és ADP arány: Az ATP a pumpa energiaforrása. Ha a sejtben az ATP szint alacsony, vagy az ADP szint magas, az enzim aktivitása csökken, mivel nincs elegendő energia a működéshez.
- pH: A sejt belső pH-ja is befolyásolhatja az enzim konformációját és aktivitását.
- Protein kinázok (PKC, PKA): A különböző protein kinázok képesek foszforilálni az Na+/K+-ATPáz alegységeit, ami megváltoztathatja az enzim aktivitását, affinitását az ionokhoz, vagy a membránba való beépülését. Például a protein kináz C (PKC) aktiválása gyakran gátolja a pumpa működését, míg a protein kináz A (PKA) aktiválása serkentheti azt.
- Oxidatív stressz: A reaktív oxigénfajták (ROS) károsíthatják az enzim fehérjeszerkezetét, csökkentve annak aktivitását, ami hozzájárulhat különböző patológiás állapotokhoz.
Ezen szabályozó mechanizmusok finomhangolása biztosítja, hogy az Na+/K+-ATPáz optimálisan működjön a sejt élettani igényeinek megfelelően, fenntartva a kritikus iongrádienseket és a sejt homeosztázisát.
Klinikai vonatkozások és patofiziológia
Az Na+/K+-ATPáz központi szerepe a sejtélettanban azt jelenti, hogy működésének zavarai számos betegség kialakulásához hozzájárulhatnak. Ezen enzimre ható gyógyszerek és toxinok pedig jelentős klinikai jelentőséggel bírnak.
Szívglikozidok (digitálisz, ouabain) hatásmechanizmusa
Talán a legismertebb klinikai vonatkozás a szívglikozidok, például a digitoxin és az ouabain hatásmechanizmusa. Ezek a vegyületek szelektíven gátolják az Na+/K+-ATPáz működését, különösen a szívizomsejtekben expresszálódó izoformákat. A gátlás eredményeként az intracelluláris nátriumkoncentráció megemelkedik. Ez az emelkedett nátriumszint csökkenti a Na+/Ca2+ kicserélő (NCX) működésének hatékonyságát, mivel az NCX a nátrium grádiensét használja fel a kalcium kiáramoltatására.
Ha az NCX kevésbé hatékony, kevesebb kalcium távozik a sejtből, ami az intracelluláris kalciumszint emelkedését eredményezi. A magasabb intracelluláris kalciumkoncentráció fokozza a szívizom kontrakciós erejét (pozitív inotrop hatás), ami a szívelégtelenség kezelésében terápiásan hasznos lehet. Ugyanakkor a túlzott gátlás aritmiákhoz és toxikus hatásokhoz vezethet, ezért a szívglikozidok adagolása precíz monitorozást igényel.
„A szívglikozidok az Na+/K+-ATPáz gátlásával fokozzák a szívizom erejét, de túladagolásuk súlyos mellékhatásokkal járhat.”
Hipertónia és az enzim
Számos kutatás vizsgálja az Na+/K+-ATPáz szerepét a hipertónia (magas vérnyomás) patogenezisében. Feltételezik, hogy az enzim aktivitásának csökkenése bizonyos egyénekben hozzájárulhat a nátrium- és vízretencióhoz, valamint az érfal simaizomsejtjeinek megnövekedett kontrakciós készségéhez. Az intracelluláris nátrium emelkedése ezekben a sejtekben szintén a Na+/Ca2+ kicserélő gátlásához vezethet, ami emelkedett intracelluláris kalciumszintet és fokozott érszűkületet okoz. Genetikai polimorfizmusok az Na+/K+-ATPáz génjeiben összefüggésbe hozhatók a magas vérnyomásra való hajlammal.
Neurológiai betegségek
Az idegsejtek rendkívül érzékenyek az iongrádiensek zavaraira, így az Na+/K+-ATPáz diszfunkciója súlyos neurológiai következményekkel járhat.
- Iszkémiás stroke: Oxigénhiányos állapotokban az ATP-termelés leáll, ami az Na+/K+-ATPáz működésének leállásához vezet. Ez az iongrádiensek összeomlását, az intracelluláris nátrium és kalcium szintjének drámai emelkedését, a sejt duzzadását és végül sejtpusztulást okoz (excitotoxicitás).
- Epilepszia: Az Na+/K+-ATPáz elégtelen működése az idegsejtek fokozott ingerlékenységéhez vezethet, hozzájárulva az epilepsziás rohamok kialakulásához. Bizonyos genetikai mutációk az enzim alegységeiben epilepsziás szindrómákhoz vezethetnek.
- Rapid-onset dystonia-parkinsonism (RDP): Ez egy ritka genetikai betegség, amelyet az ATP1A3 gén mutációja okoz, amely az Na+/K+-ATPáz α3 alegységét kódolja. A betegség hirtelen fellépő dystoniával és Parkinson-szerű tünetekkel jár, rávilágítva az enzim kulcsszerepére az idegrendszer normális működésében.
Vesebetegségek
Mint már említettük, a vese működése nagymértékben függ az Na+/K+-ATPáz aktivitásától. Az enzim diszfunkciója vesebetegségekhez, elektrolit-egyensúly zavarokhoz (pl. hyperkalémia, hyponatrémia) és folyadékretencióhoz vezethet. Bizonyos vesebetegségekben az enzim expressziója vagy aktivitása megváltozhat, hozzájárulva a patológiás folyamatokhoz.
Rák és az Na+/K+-ATPáz
Az elmúlt évek kutatásai rámutattak az Na+/K+-ATPáz potenciális szerepére a rák kialakulásában és progressziójában. A daganatsejtek gyakran eltérő módon expresszálják az enzim izoformáit, vagy megváltozik az aktivitásuk. Bizonyos esetekben az enzim gátlása (pl. szívglikozidokkal) antiproliferatív és apoptózist indukáló hatást mutatott daganatsejtekben, ami új terápiás lehetőségeket nyithat meg. Az enzim membránban lévő lokalizációja és jelátviteli partnerekkel való interakciója is befolyásolhatja a tumorsejt-biológiát.
Cisztás fibrózis
Bár az Na+/K+-ATPáz nem közvetlenül érintett a cisztás fibrózis (CF) alaphibájában (CFTR gén mutációja), a CFTR csatorna diszfunkciója által okozott ion- és folyadéktranszport zavarok indirekt módon befolyásolhatják az Na+/K+-ATPáz aktivitását. A CF-ben szenvedő betegek légúti hámsejtjeiben megváltozik az iontranszport, ami a mucociliáris clearance károsodásához és krónikus fertőzésekhez vezet. Az ioncsatornák és pumpák, köztük az Na+/K+-ATPáz közötti komplex kölcsönhatás kritikus a légutak folyadékegyensúlyának fenntartásában.
Összességében az Na+/K+-ATPáz diszfunkciója számos betegség alapját képezheti, vagy hozzájárulhat azok progressziójához. Az enzim mechanizmusának és szabályozásának mélyebb megértése új terápiás stratégiák kidolgozását teszi lehetővé ezeknek a komplex állapotoknak a kezelésére.
Az Na+/K+-ATPáz mint terápiás célpont

Az Na+/K+-ATPáz létfontosságú szerepe és számos patofiziológiai folyamatban való érintettsége miatt kiváló terápiás célpontot jelent a gyógyszerfejlesztés számára. A már említett szívglikozidok, mint a digitálisz, évtizedek óta használatosak a szívelégtelenség kezelésében, kifejezetten az enzim gátlásán keresztül fejtve ki hatásukat. Azonban a szívglikozidok szűk terápiás ablaka és számos mellékhatása arra ösztönzi a kutatókat, hogy új, specifikusabb és biztonságosabb molekulákat keressenek.
A modern gyógyszerkutatás egyik iránya az izoform-specifikus gátlók vagy aktivátorok fejlesztése. Mivel az Na+/K+-ATPáz különböző α-alegységei (α1, α2, α3, α4) eltérő szöveti eloszlással és funkciókkal rendelkeznek, egy olyan gyógyszer, amely szelektíven célozná például az α2 izoformát (gyakori a szívben és az izmokban) az α1 izoformát érintetlenül hagyva (mindenütt jelen van), sokkal specifikusabb terápiás hatást érhetne el kevesebb mellékhatással. Ez különösen releváns lehet a szívbetegségek, neurológiai rendellenességek vagy akár a rák kezelésében.
Emellett folynak kutatások az Na+/K+-ATPáz aktivátorainak azonosítására is. Bizonyos állapotokban, például hypokalemia vagy az iontranszport károsodása esetén, az enzim aktivitásának fokozása előnyös lehet. Az ilyen aktivátorok segíthetnek helyreállítani az ionegyensúlyt és javítani a sejtműködést.
Az enzim jelátviteli szerepének feltárása is új terápiás utakat nyithat meg. Az Na+/K+-ATPáz nem csupán egy ionpumpa, hanem egy jelátviteli receptor is, amely képes kölcsönhatásba lépni más fehérjékkel, és intracelluláris jelátviteli útvonalakat aktiválni. Ezen jelátviteli funkciók modulálása, anélkül, hogy az iontranszportot befolyásolnánk, szintén ígéretes terápiás stratégia lehetne. Ez a megközelítés különösen érdekes a daganatos megbetegedések és a gyulladásos folyamatok kontextusában, ahol az enzim jelátviteli szerepe patológiásan megváltozhat.
A Na+/K+-ATPáz kutatása továbbra is intenzíven zajlik, a szerkezeti biológiai elemzésektől a preklinikai és klinikai vizsgálatokig. A cél az, hogy jobban megértsük ennek a fundamentális enzimnek a komplex működését és szabályozását, és ezen ismeretek alapján új, hatékonyabb és biztonságosabb gyógyszereket fejlesszünk ki a súlyos betegségek kezelésére.
Kutatási irányok és jövőbeli perspektívák
Az Na+/K+-ATPáz, mint a sejtélettan egyik sarokköve, továbbra is a modern biológia és orvostudomány kiemelt kutatási területe. A jövőbeli kutatások számos izgalmas irányba mutatnak, amelyek mélyebb betekintést nyújtanak az enzim működésébe és potenciális terápiás alkalmazásaiba.
Szerkezeti biológia és mechanizmus finomhangolása
A krioszintetikus elektronmikroszkópia (cryo-EM) és más nagy felbontású szerkezeti módszerek fejlődése lehetővé teszi az Na+/K+-ATPáz különböző konformációs állapotainak atomi szintű vizsgálatát. Ennek köszönhetően egyre pontosabb képet kapunk az ionkötő helyekről, az ATP hidrolízisének dinamikájáról és a konformációs változások molekuláris mechanizmusairól. A jövőbeli kutatások valószínűleg a különböző izoformák és a γ-alegységek pontos szerepére fókuszálnak majd a szerkezet-funkció összefüggésben, ami elengedhetetlen a specifikus gyógyszerek tervezéséhez.
Jelátviteli szerep és interaktóm
Egyre világosabbá válik, hogy az Na+/K+-ATPáz nem csupán egy passzív pumpa, hanem aktív szereplő a sejten belüli jelátviteli hálózatokban. Interagál más fehérjékkel, például receptorokkal, kinázokkal és scaffold fehérjékkel, és képes aktiválni különböző jelátviteli útvonalakat (pl. MAPK, Src kináz). A jövőbeli kutatások célja az Na+/K+-ATPáz interaktómjának (az összes vele kölcsönható fehérje) teljes feltérképezése, valamint ezen jelátviteli funkciók élettani és patofiziológiai szerepének tisztázása. Ez újabb terápiás célpontokat azonosíthat, amelyek az iontranszport befolyása nélkül modulálnák a sejtfunkciókat.
Betegségek specifikus izoformáinak vizsgálata
Ahogy a genetikai vizsgálatok egyre hozzáférhetőbbé válnak, egyre több betegséget hoznak összefüggésbe az Na+/K+-ATPáz különböző izoformáinak mutációival. A Rapid-onset dystonia-parkinsonism (ATP1A3), a familiáris hemiplegiás migrén (ATP1A2) és más neurológiai zavarok (pl. epilepszia, autizmus spektrumzavarok) mind az enzim genetikai defektusaihoz köthetők. A jövőben részletesebben vizsgálják majd, hogy ezek a mutációk pontosan hogyan befolyásolják az enzim működését, és hogyan lehetne ezeket a specifikus hibákat célzottan korrigálni génterápiás vagy farmakológiai megközelítésekkel.
Az Na+/K+-ATPáz szerepe az öregedésben és neurodegenerációban
Az Na+/K+-ATPáz aktivitása csökkenhet az öregedés során, ami hozzájárulhat a sejtek ionhomeosztázisának romlásához és a neurodegeneratív folyamatokhoz. Kutatások vizsgálják, hogy az enzim aktivitásának fenntartása vagy fokozása lassíthatja-e az öregedési folyamatokat, vagy védelmet nyújthat-e az Alzheimer-kór, Parkinson-kór és más neurodegeneratív betegségek ellen. Az oxidatív stressz és a mitokondriális diszfunkció hatása az enzimre szintén fontos kutatási terület.
Új terápiás stratégiák és gyógyszerek
A fenti kutatási irányok mind a jövőbeli terápiás stratégiák alapjait fektetik le. Az izoform-specifikus gátlók és aktivátorok, a jelátviteli funkciókat moduláló vegyületek, valamint a génterápiás megközelítések mind ígéretes lehetőségeket kínálnak. A mesterséges intelligencia és a gépi tanulás alkalmazása a gyógyszertervezésben felgyorsíthatja az új, nagy affinitású és szelektivitású molekulák azonosítását, amelyek az Na+/K+-ATPáz különböző aspektusait célozzák.
Összességében az Na+/K+-ATPáz továbbra is a biológiai kutatás élvonalában marad, és a róla szerzett új ismeretek forradalmasíthatják a betegségek megértését és kezelését a sejtélettan, a neurológia, a kardiológia és az onkológia területén egyaránt.







