
Az élet alapvető építőkövei a fehérjék, amelyek sejteink működésének szinte minden aspektusában kulcsszerepet játszanak. Gondoljunk csak az enzimekre, amelyek felgyorsítják a kémiai reakciókat, a hormonokra, melyek üzeneteket közvetítenek, vagy az antitestekre, amelyek megvédenek minket a kórokozóktól. Ezek a komplex molekulák azonban nem léteznének a fehérjeszintézis, azaz a fehérjék előállításának csodálatos és precíz folyamata nélkül. Ez a biológiai mechanizmus minden élő szervezetben zajlik, a legegyszerűbb baktériumoktól kezdve egészen az emberi test milliárdnyi sejtjéig, biztosítva az élet fenntartásához szükséges folyamatos megújulást és alkalmazkodást. A génexpresszió központi elemeként a fehérjeszintézis az a híd, amely a genetikai információt a működőképes biológiai struktúrákká alakítja.
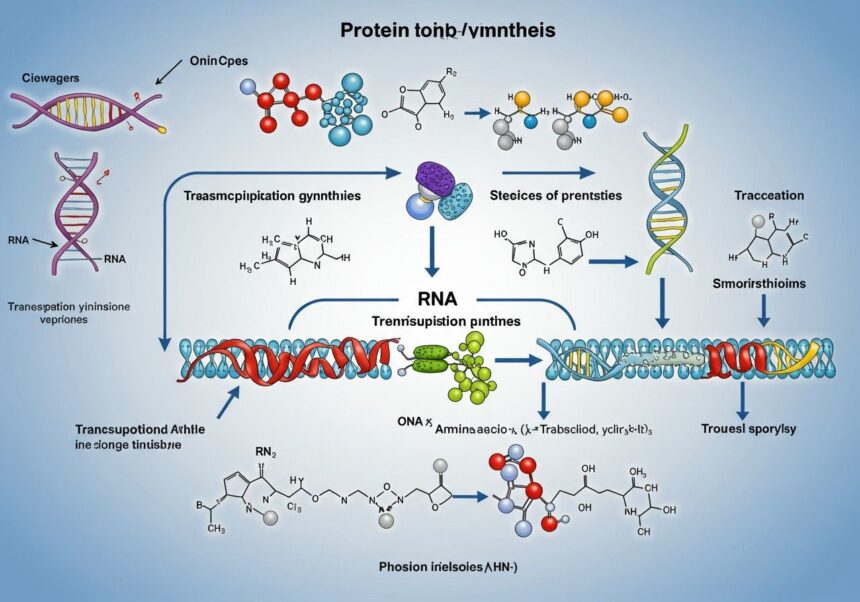
A fehérjeszintézis egy olyan bonyolult, mégis rendkívül szervezett folyamat, amely során a sejtek a DNS-ben tárolt genetikai információt felhasználva, lépésről lépésre felépítik az aminosavakból álló fehérjemolekulákat. Ez a mechanizmus nem csupán az új fehérjék előállítását jelenti, hanem a sérült vagy elöregedett fehérjék pótlását is, biztosítva a sejtek és szövetek dinamikus egyensúlyát. Az egész folyamat két fő szakaszra bontható: a transzkripcióra (átírásra) és a transzlációra (fordításra). E két fázis során a genetikai kód „leolvasásra” és „lefordításra” kerül, hogy a sejt pontosan a megfelelő fehérjét állítsa elő a megfelelő időben és mennyiségben.
Ahhoz, hogy megértsük a fehérjeszintézis alapjait, először ismernünk kell a főszereplőket. A genetikai információ tárolója a DNS, amely a sejtmagban található, és kettős spirál formájában hordozza az összes örökletes utasítást. A fehérjeszintézishez azonban a DNS-nek nem kell elhagynia a sejtmag biztonságát. Ehelyett egy közvetítő molekula, az RNS (ribonukleinsav) viszi át az információt a sejtmagból a citoplazmába, ahol a tényleges fehérjeépítés zajlik. Különböző típusú RNS-ek vesznek részt ebben a folyamatban, mindegyiknek megvan a maga specifikus feladata, biztosítva a szintézis zökkenőmentes lefolyását.
A fehérjeszintézis alapjai és jelentősége
A fehérjeszintézis, más néven proteinszintézis, az a biológiai folyamat, amely során a sejtek aminosavakból polipeptid láncokat, majd ebből funkcionális fehérjéket hoznak létre. Ez a folyamat a centrális dogma része, amely kimondja, hogy a genetikai információ a DNS-ből az RNS-en keresztül áramlik a fehérjékbe. Ez a dogma alapvető fontosságú az élet megértéséhez, mivel magyarázza, hogyan öröklődnek és fejeződnek ki a genetikai tulajdonságok.
A fehérjék szerepe a szervezetben rendkívül sokrétű és nélkülözhetetlen. Képzeljük el a sejteket, mint egy összetett gépezetet, ahol a fehérjék a gépezet működésének minden részletét irányítják. Az enzimek például fehérjék, amelyek biokémiai reakciókat katalizálnak, felgyorsítva azokat a sebességet, ami nélkül az életfolyamatok nem mennének végbe. Gondoljunk csak az emésztésre, ahol az enzimek bontják le az élelmiszert, vagy a légzésre, ahol energiát termelnek a tápanyagokból. Nélkülük a sejtanyagcsere megállna.
A fehérjék emellett szerkezeti elemekként is funkcionálnak. A kollagén és az elasztin például a bőr, a csontok és a kötőszövetek fő alkotóelemei, rugalmasságot és szilárdságot biztosítva. Az aktin és a miozin izmaink összehúzódásáért felelős fehérjék, lehetővé téve a mozgást. Ezek a szerkezeti fehérjék biztosítják a sejtek és szövetek fizikai integritását és formáját.
A transzportfehérjék, mint például a hemoglobin, oxigént szállítanak a vérben, míg más fehérjék a sejtmembránon keresztül juttatják be vagy ki a tápanyagokat és salakanyagokat. Az immunrendszerünk alapját képező antitestek is fehérjék, amelyek felismerik és semlegesítik a kórokozókat, védelmet nyújtva a betegségekkel szemben. Ezenkívül számos hormon, mint például az inzulin, szintén fehérje természetű, és a szervezet különböző funkcióit szabályozza, mint például a vércukorszintet.
„A fehérjeszintézis nem csupán egy biokémiai folyamat, hanem az életfolyamatok karmestere, amely a genetikai kód dallamát funkcionális harmóniává alakítja.”
A sejtekben a fehérjeszintézis fő helyszínei a riboszómák. Ezek a komplex molekuláris gépezetek felelősek az mRNS-ben kódolt információ lefordításáért. Eukarióta sejtekben a riboszómák szabadon lebeghetnek a citoplazmában, vagy kapcsolódhatnak az endoplazmatikus retikulumhoz (ER), egy membránrendszerhez, amely a sejten belüli anyagok szállításáért és feldolgozásáért felelős. A sejtmagban zajlik a transzkripció, ahol a DNS-ről RNS másolat készül, míg a transzláció, a tényleges fehérjeépítés, a citoplazmában, a riboszómákon történik.
A DNS (dezoxiribonukleinsav) a genetikai információ archívuma. Kettős spirál alakú molekula, amely négyféle nukleotidból épül fel: adenin (A), guanin (G), citozin (C) és timin (T). Ezek a bázisok specifikus párokban kapcsolódnak egymáshoz (A-T, G-C), és a sorrendjük hordozza a genetikai kódot. A gén a DNS egy olyan szakasza, amely egy adott fehérje vagy RNS molekula szintézisére vonatkozó információt tartalmazza.
Az RNS (ribonukleinsav) a DNS-hez hasonló, de egyszálú molekula, amely timin helyett uracilt (U) tartalmaz. Három fő típusa vesz részt a fehérjeszintézisben:
- mRNS (messenger RNS): Ez a hírvivő molekula, amely a DNS-ről másolt genetikai információt szállítja a riboszómákhoz. Az mRNS sorrendje határozza meg az aminosavak sorrendjét a fehérjében.
- tRNS (transzfer RNS): Ez a „fordító” molekula. Minden tRNS egy specifikus aminosavat szállít, és egy antikodon nevű szekvenciával rendelkezik, amely képes párosodni az mRNS-en lévő kodonokkal.
- rRNS (riboszomális RNS): Ez az RNS-típus a riboszómák fő alkotóeleme, és katalitikus szerepet játszik a peptidkötések kialakításában az aminosavak között.
Az aminosavak a fehérjék építőkövei. Összesen 20 különböző aminosav létezik, amelyek a peptidkötések révén kapcsolódnak egymáshoz, hosszú láncokat, azaz polipeptideket alkotva. A polipeptid láncok feltekeredve és specifikus térszerkezetet felvéve válnak funkcionális fehérjékké. Az aminosavak sorrendje határozza meg a fehérje egyedi térszerkezetét és funkcióját.
A genetikai kód az a szabályrendszer, amely meghatározza, hogy az mRNS nukleotidjainak sorrendje hogyan fordítódik le aminosav sorrenddé. A kód hármas egységekből, úgynevezett kodonokból áll, amelyek mindegyike egy specifikus aminosavat kódol. Például az AUG kodon a metionint kódolja, és egyben a start kodonként is funkcionál, jelezve a fehérjeszintézis kezdetét. A genetikai kód degenerált, ami azt jelenti, hogy több kodon is kódolhatja ugyanazt az aminosavat. Ez a „redundancia” bizonyos mértékig védelmet nyújt a mutációk káros hatásai ellen. A kód emellett szinte univerzális, vagyis ugyanazok a kodonok ugyanazokat az aminosavakat kódolják a baktériumoktól az emberig, ami az élet közös eredetére utal.
Az első lépés: a transzkripció (átírás)
A transzkripció az a folyamat, amely során a DNS-ben tárolt genetikai információ egy RNS molekulába másolódik át. Ez a lépés alapvető a génexpresszió szempontjából, mivel a DNS molekula túl nagy és túl értékes ahhoz, hogy elhagyja a sejtmag védelmét. Ehelyett egy kisebb, mobilisabb RNS másolat, az mRNS készül, amely képes eljutni a citoplazmába a riboszómákhoz.
A transzkripció helyszíne eltérő az élőlénytípusok között. Eukarióta sejtekben, mint például az emberi sejtekben, a transzkripció a sejtmagban zajlik, ahol a DNS található. Az elkészült mRNS ezután a sejtmagpórusokon keresztül jut ki a citoplazmába. Prokarióta sejtekben, mint például a baktériumokban, amelyek nem rendelkeznek sejtmaggal, a transzkripció és a transzláció a citoplazmában történik, sőt, gyakran szinte egyidejűleg.
A folyamat kezdetén a DNS kettős spiráljának egy szakasza, a gén, szolgál templátként. A DNS szálai szétválnak, és az egyik szál (a templát szál) nukleotid sorrendje alapján épül fel az új RNS molekula. A DNS templát bázisaihoz komplementer RNS bázisok kapcsolódnak: guaninhoz citozin, citozinhoz guanin, adeninhez uracil (a timin helyett), és timinhez adenin. Ez biztosítja az információ pontos átvitelét.
A transzkripció kulcsenzime az RNS-polimeráz. Ez az enzim végzi a DNS szálainak szétválasztását, az új RNS nukleotidok egymáshoz kapcsolását, és az RNS lánc növelését. Az RNS-polimeráz nem igényel primert a szintézis megkezdéséhez, ellentétben a DNS-polimerázzal.
A transzkripció három fő fázisra osztható:
- Iniciáció (kezdés): Az RNS-polimeráz felismeri és hozzákötődik a DNS egy specifikus régiójához, az úgynevezett promóterhez. A promóterek olyan szabályozó szekvenciák, amelyek jelzik az RNS-polimeráznak, hogy hol kezdje meg a transzkripciót, és melyik DNS szálat használja templátként. Eukariótákban számos transzkripciós faktor segít az RNS-polimeráznak a promóter felismerésében és a kötődésben, szabályozva ezzel a génexpressziót. A DNS kettős spirálja feltekeredik, és egy transzkripciós buborék alakul ki.
- Elongáció (lánchosszabbítás): Az RNS-polimeráz elindul a templát szál mentén, és sorban hozzáadja a komplementer RNS nukleotidokat, miközben az RNS lánc folyamatosan növekszik. Az enzim haladása során a DNS kettős spirálja előtte szétválik, mögötte pedig újra összezáródik. Az RNS-polimeráz nagy pontossággal dolgozik, de hibaaránya magasabb, mint a DNS-polimerázé, mivel az RNS molekulák kevésbé stabilak és kevésbé hosszú életűek, mint a DNS.
- Termináció (befejezés): A transzkripció akkor fejeződik be, amikor az RNS-polimeráz elér egy specifikus DNS szekvenciát, az úgynevezett terminátor régiót. A terminátor jelzi az enzimnek, hogy váljon le a DNS-ről, és engedje el az újonnan szintetizált RNS molekulát. Prokariótákban gyakran közvetlenül a riboszómákhoz kapcsolódhat a frissen szintetizált mRNS, míg eukariótákban további feldolgozásra van szükség.
Eukarióta sejtekben az újonnan szintetizált RNS molekula, a pre-mRNS, még nem teljesen érett és funkcionális. Számos módosításon kell átesnie, mielőtt elhagyhatja a sejtmagot és részt vehet a transzlációban. Ezeket a módosításokat poszt-transzkripciós módosításoknak nevezzük, és kulcsfontosságúak az mRNS stabilitásához, szállításához és a hatékony transzlációhoz.
A legfontosabb poszt-transzkripciós módosítások a következők:
- 5′ sapkázás (5′ cap): Az mRNS 5′ végére egy speciális nukleotid, a 7-metilguanozin kerül. Ez a „sapka” védi az mRNS-t a lebontástól, segíti a sejtmagból való kijutását, és fontos szerepet játszik a riboszómák általi felismerésben a transzláció kezdetén.
- 3′ poli-A farok (3′ poly-A tail): Az mRNS 3′ végére számos adenin nukleotidból álló lánc, a poli-A farok kapcsolódik. Ez a farok szintén védi az mRNS-t a lebontástól, és befolyásolja az mRNS élettartamát a citoplazmában. Minél hosszabb a poli-A farok, annál tovább él az mRNS.
- Splicing (összeillesztés): Ez az egyik legkomplexebb és legfontosabb módosítás. Az eukarióta gének gyakran tartalmaznak kódoló (exon) és nem kódoló (intron) szakaszokat. Az intronokat el kell távolítani a pre-mRNS-ből, és az exonokat pontosan össze kell illeszteni, hogy egy folytonos, kódoló mRNS lánc jöjjön létre. Ezt a folyamatot a spliceoszóma nevű nagy RNS-protein komplex végzi.
A splicing különösen érdekes aspektusa az alternatív splicing. Ez azt jelenti, hogy egyetlen gén pre-mRNS-éből különböző exon kombinációkkal több különböző mRNS molekula is létrejöhet. Ennek eredményeként egyetlen gén számos különböző fehérjét kódolhat, jelentősen növelve a genom komplexitását és a fehérjék sokféleségét. Ez magyarázza, hogy miért van viszonylag kevés génünk (körülbelül 20 000-25 000), mégis sokkal több fehérjénk.
Miután az összes poszt-transzkripciós módosítás megtörtént, az érett mRNS készen áll arra, hogy elhagyja a sejtmagot, és elinduljon a citoplazmába, ahol a következő lépés, a transzláció vár rá. Az mRNS stabilitása és élettartama alapvető a génexpresszió szabályozásában, mivel befolyásolja, hogy mennyi ideig és milyen gyakran fordítódik le egy adott fehérje.
A második lépés: a transzláció (fordítás)
A transzláció, vagyis a fordítás, a fehérjeszintézis második fő szakasza, amely során az mRNS-ben tárolt genetikai információ aminosav sorrenddé, azaz egy polipeptid lánccá alakul. Ez a folyamat a sejt „gyártósorán”, a riboszómákon zajlik, amelyek a citoplazmában találhatók. Eukarióta sejtekben a riboszómák szabadon úszhatnak a citoplazmában, vagy kapcsolódhatnak az endoplazmatikus retikulumhoz (ER). Az ER-hez kötött riboszómák általában olyan fehérjéket szintetizálnak, amelyek a sejtmembránba épülnek, a sejtből kiválasztódnak, vagy a lizoszómákba, illetve más organellumokba kerülnek.
A mRNS a „receptkönyv”, amely a fehérje felépítéséhez szükséges utasításokat tartalmazza. Az mRNS molekula nukleotidjainak sorrendje határozza meg az aminosavak pontos sorrendjét a készülő fehérjében. A genetikai kód hármas egységekben, úgynevezett kodonokban van leírva, ahol minden kodon egy specifikus aminosavat kódol, vagy jelzi a szintézis befejezését (stop kodon).
A tRNS (transzfer RNS) molekulák a „fordítók” vagy „szállítók”. Minden tRNS molekula két fontos régióval rendelkezik: az egyik végén egy specifikus aminosav kötődik hozzá, a másik végén pedig egy három nukleotidból álló szekvencia található, az antikodon. Az antikodon képes komplementer módon párosodni az mRNS-en lévő kodonnal. Például, ha az mRNS-en az UUU kodon található, akkor a tRNS-en az AAA antikodon fog ehhez kötődni, és ez a tRNS szállítja a fenilalanin aminosavat. A tRNS molekulák aminosavakkal való feltöltését speciális enzimek, az aminosav-tRNS szintetázok végzik, rendkívüli pontossággal.
A riboszóma a transzláció tényleges „gyártósori gépezete”. Két alegységből áll: egy nagy és egy kis alegységből. Ezek az alegységek rRNS (riboszomális RNS) és riboszomális fehérjék komplexéből épülnek fel. A riboszóma feladata az mRNS „leolvasása” és a megfelelő tRNS-ek beillesztése, valamint a peptidkötések kialakítása az aminosavak között. A riboszómán belül három kulcsfontosságú hely található:
- A-hely (aminoacil-tRNS kötőhely): Ide érkezik az új, aminosavval feltöltött tRNS molekula, amelynek antikodonja párosodik az mRNS kodonjával.
- P-hely (peptidil-tRNS kötőhely): Itt található az a tRNS, amely a növekvő polipeptid láncot hordozza.
- E-hely (kilépési hely): Innen távozik a riboszómáról az aminosavját már leadott tRNS.
A transzláció is három fő fázisra osztható:
- Iniciáció (kezdés): A transzláció a start kodon (általában AUG) felismerésével kezdődik az mRNS-en. Eukariótákban az iniciátor tRNS, amely a metionin aminosavat szállítja, először a kis riboszóma alegységhez kötődik. Ez a komplex ezután végigsiklik az mRNS-en, amíg el nem éri az AUG start kodont. Amint megtalálja, a nagy riboszóma alegység is csatlakozik, és a teljes riboszóma összeáll a start kodon körül. A metionin tRNS ekkor a P-helyen helyezkedik el.
- Elongáció (lánchosszabbítás): Ez a fázis az aminosavak egymás utáni hozzáadását jelenti a növekvő polipeptid lánchoz.
- Kodon-antikodon párosítás: Egy új, aminosavval feltöltött tRNS érkezik az A-helyre, amelynek antikodonja komplementer az mRNS-en lévő kodonnal.
- Peptidkötés képződése: A riboszóma rRNS-e (amely ribozimként működik) katalizálja a peptidkötés kialakulását a P-helyen lévő tRNS által hordozott polipeptid lánc és az A-helyen lévő új aminosav között. Ekkor a polipeptid lánc átkerül az A-helyen lévő tRNS-re.
- Transzlokáció: A riboszóma pontosan egy kodonnyit elmozdul az mRNS mentén 5′ irányból 3′ irányba. Ennek hatására az A-helyen lévő tRNS a P-helyre kerül (most már a teljes polipeptid lánccal), a P-helyen lévő, üres tRNS az E-helyre kerül, ahonnan kilép a riboszómáról, az A-hely pedig szabaddá válik a következő aminosav-tRNS számára. Ez a ciklus ismétlődik, amíg a polipeptid lánc teljesen fel nem épül.
- Termináció (befejezés): Az elongáció akkor ér véget, amikor a riboszóma elér egy stop kodont az mRNS-en (UAA, UAG, vagy UGA). Ezek a kodonok nem kódolnak aminosavat. Ehelyett felszabadító faktorok (release factors) kötődnek a stop kodonhoz az A-helyen. Ezek a faktorok elősegítik a polipeptid lánc leválását a tRNS-ről és a riboszómáról. Ezt követően a riboszóma alegységei szétválnak, és az mRNS is felszabadul.
A hatékony fehérjeszintézis érdekében gyakran több riboszóma is képes egyszerre leolvasni ugyanazt az mRNS molekulát. Ezt a struktúrát poliszómának vagy poliriboszómának nevezik. A poliszómák lehetővé teszik, hogy egyetlen mRNS molekuláról egyszerre több fehérjemolekula is szintetizálódjon, ami jelentősen növeli a fehérjetermelés sebességét és hatékonyságát.
A transzláció rendkívül komplex és finoman szabályozott folyamat, amely a sejt energiafelhasználásának jelentős részét teszi ki. A pontosság elengedhetetlen, hiszen egyetlen tévesen beépített aminosav is megváltoztathatja a fehérje térszerkezetét és funkcióját, súlyos következményekkel járva a sejt és az egész szervezet számára.
A fehérjék útja a szintézistől a funkcióig
A fehérjeszintézis befejezésével, amikor a polipeptid lánc levált a riboszómáról, a munka még korántsem ért véget. Ahhoz, hogy egy polipeptid lánc funkcionális fehérjévé váljon, számos utólagos módosításon és folyamaton kell átesnie. Ezeket poszt-transzlációs módosításoknak nevezzük, és kulcsfontosságúak a fehérje megfelelő térszerkezetének kialakításában, célba juttatásában és szabályozásában.
Az egyik legfontosabb poszt-transzlációs lépés a fehérjehajtogatás (protein folding). A frissen szintetizált polipeptid lánc lineáris aminosav sorrendje az elsődleges szerkezet. Ahhoz, hogy funkcionális legyen, ez a lánc specifikus és komplex háromdimenziós térszerkezetet kell, hogy felvegyen. Ez a folyamat nagyrészt spontán módon, az aminosavak közötti kölcsönhatások (hidrogénkötések, diszulfidhidak, hidrofób interakciók) révén megy végbe, de gyakran igényel segítséget.
Ebben a folyamatban kulcsszerepet játszanak a chaperonok (hőstressz-fehérjék). Ezek a segítő fehérjék megakadályozzák a polipeptid láncok nem megfelelő aggregálódását vagy téves hajtogatását, különösen stresszhelyzetben, például magas hőmérsékleten. A chaperonok segítenek a polipeptid láncnak megtalálni a legstabilabb és funkcionális térszerkezetét. A tévesen hajtogatott fehérjék súlyos problémákat okozhatnak, mivel hajlamosak aggregálódni, ami számos neurodegeneratív betegség, például az Alzheimer-kór, Parkinson-kór vagy a prion betegségek alapját képezi.
A fehérjék térszerkezete négy hierarchikus szinten írható le:
- Elsődleges szerkezet: Az aminosavak lineáris sorrendje.
- Másodlagos szerkezet: Helyi, ismétlődő mintázatok, mint az alfa-hélixek és béta-redők, amelyeket hidrogénkötések stabilizálnak.
- Harmadlagos szerkezet: Az egész polipeptid lánc háromdimenziós alakja, amely a másodlagos szerkezeti elemek további hajtogatásával jön létre. Ez az alak határozza meg a fehérje funkcióját.
- Negyedleges szerkezet: Ha több polipeptid lánc (alegység) kapcsolódik egymáshoz, hogy egy funkcionális fehérjekomplexet alkosson, akkor beszélünk negyedleges szerkezetről (pl. hemoglobin).
A hajtogatáson kívül számos kovalens módosítás is történhet a fehérjéken, amelyek jelentősen befolyásolhatják azok aktivitását, stabilitását, lokalizációját és kölcsönhatásait. Néhány példa:
- Foszforiláció: Foszfátcsoport hozzáadása egy aminosavhoz (általában szerinhez, treoninhoz vagy tirozinhoz). Ez az egyik leggyakoribb szabályozó mechanizmus, amely ki- vagy bekapcsolhatja a fehérje aktivitását, jelezve a sejtnek a változó körülményeket.
- Glikoziláció: Szénhidrátláncok hozzáadása a fehérjéhez. Ez gyakran a sejtfelszíni fehérjéknél vagy a kiválasztott fehérjéknél fordul elő, és szerepet játszik a sejt-sejt felismerésben és az immunválaszban.
- Acetiláció és metiláció: Acetil- vagy metilcsoportok hozzáadása, amelyek befolyásolhatják a fehérje stabilitását, kölcsönhatásait vagy a génexpressziót (különösen a hiszton fehérjéknél).
- Proteolitikus hasítás: A polipeptid lánc bizonyos szakaszainak eltávolítása enzimek (proteázok) által. Ez gyakran inaktív prekurzor fehérjék (proenzimek, prohormonok) aktiválásához vezet, például az inzulin vagy emésztőenzimek esetében.
- Diszulfidhidak képződése: Két cisztein aminosav tiolcsoportja közötti kovalens kötés, amely stabilizálja a fehérje térszerkezetét, különösen az extrazelluláris fehérjéknél.
A fehérjéknek nemcsak megfelelő térszerkezetet kell felvenniük, hanem a sejtben a megfelelő helyre is kell jutniuk ahhoz, hogy funkcionálhassanak. Ezt a folyamatot fehérje célba juttatásnak (protein targeting) nevezzük. A frissen szintetizált fehérjék egy része a citoplazmában marad, mások azonban speciális organellumokba (pl. mitokondriumok, kloroplasztiszok, peroxiszómák) vagy a sejtből kifelé (szekréció) kerülnek. Az endoplazmatikus retikulumhoz (ER) kötött riboszómákon szintetizálódó fehérjék gyakran egy szignálpeptidet tartalmaznak a N-terminális végükön, amely irányítja őket az ER membránjához. Az ER-be belépve a fehérjék további módosításokon eshetnek át, majd a Golgi-készülékbe kerülhetnek, ahonnan szétoszlanak a sejt különböző részeibe, vagy a sejtmembránba épülnek, illetve kiválasztódnak.
A sejt rendkívül szigorú minőség-ellenőrzési mechanizmusokkal rendelkezik a fehérjék hajtogatására vonatkozóan. A tévesen hajtogatott vagy sérült fehérjéket a sejt felismeri és lebontja. Ennek kulcsfontosságú rendszere az ubikvitin-proteaszóma rendszer. Az ubikvitin egy kis fehérje, amely kovalensen kapcsolódik a lebontásra szánt fehérjékhez, mintegy „címkézve” azokat. Ezek a címkézett fehérjék ezután egy nagy fehérjekomplexbe, a proteaszómába kerülnek, ahol lebontódnak aminosavakra, amelyek újra felhasználhatók a fehérjeszintézisben. Ez a folyamat elengedhetetlen a sejtek egészségének fenntartásához és a fehérjék homeosztázisának szabályozásához.
„A fehérjék, mint a molekuláris világ műalkotásai, nem csak felépülnek, hanem gondosan formálódnak, irányítódnak és szabályozódnak, hogy tökéletesen illeszkedjenek a sejt komplex működésébe.”
Összességében a fehérjék útja a genetikai információtól a funkcionális molekuláig egy rendkívül komplex és precíz folyamat, amely magában foglalja a transzkripciót, transzlációt, hajtogatást, poszt-transzlációs módosításokat és a célba juttatást. Ezen lépések bármelyikének zavara súlyos következményekkel járhat, és számos betegség hátterében állhat.
A génexpresszió szabályozása: amikor a sejt dönt
A fehérjeszintézis nem egy állandóan maximális sebességgel zajló, gátlástalan folyamat. Éppen ellenkezőleg, a sejtek rendkívül finoman hangolt mechanizmusokkal szabályozzák, hogy mely génekről készüljön RNS másolat, és mely RNS-ek fordítódjanak le fehérjévé, mikor és milyen mennyiségben. Ezt a komplex szabályozó hálózatot nevezzük génexpresszió szabályozásnak. Ennek fontossága óriási, hiszen ez teszi lehetővé a sejtek specializációját, az alkalmazkodást a változó környezeti feltételekhez, és az energia takarékos felhasználását.
Miért olyan kulcsfontosságú ez a szabályozás? Először is, a különböző sejttípusoknak más és más fehérjékre van szükségük. Egy idegsejtnek teljesen más fehérjekészlete van, mint egy izomsejtnek vagy egy májsejtnek, annak ellenére, hogy mindegyik sejt ugyanazt a genetikai információt tartalmazza (ugyanazt a DNS-t). A génexpresszió szabályozása teszi lehetővé ezt a specializációt, biztosítva, hogy minden sejt csak azokat a géneket fejezze ki, amelyekre a funkciójához szüksége van.
Másodszor, a sejteknek folyamatosan alkalmazkodniuk kell a környezeti változásokhoz. Ha például egy baktérium laktózt tartalmazó környezetbe kerül, be kell kapcsolnia azokat a géneket, amelyek a laktóz lebontásához szükséges enzimeket kódolják. Ha a laktóz eltűnik, ezeket a géneket ki kell kapcsolni, hogy energiát takarítson meg. Ez a dinamikus válasz a környezeti ingerekre szintén a génexpresszió szabályozásán keresztül valósul meg.
Harmadszor, a fehérjeszintézis energetikailag rendkívül költséges folyamat. A felesleges fehérjék előállítása pazarolja a sejt erőforrásait. A szabályozás lehetővé teszi, hogy a sejt csak azokat a fehérjéket szintetizálja, amelyekre valóban szüksége van, ezzel optimalizálva az energiafelhasználást.
A génexpresszió szabályozása több szinten is megvalósulhat, a DNS-től egészen a kész fehérje poszt-transzlációs módosításáig:
- Transzkripciós szintű szabályozás: Ez a leggyakoribb és legfontosabb szabályozási szint. A sejt itt dönti el, hogy egyáltalán elkészüljön-e az RNS másolat egy adott génről. Ennek mechanizmusai a következők:
- Promóterek és enhancerek: A promóterekhez kötődik az RNS-polimeráz, és jelzik a transzkripció kezdetét. Az enhancerek távolabb elhelyezkedő DNS-régiók, amelyekhez specifikus fehérjék (aktivátorok) kötődnek, és felerősítik a transzkripciót.
- Represszorok és aktivátorok: A represszor fehérjék a promóterhez vagy a promóter közelében lévő operátor régióhoz kötődve gátolják az RNS-polimeráz működését, megakadályozva a transzkripciót. Az aktivátor fehérjék viszont segítik az RNS-polimeráz kötődését és a transzkripció elindítását.
- Kromatin szerkezet módosítása: Eukariótákban a DNS hiszton fehérjék köré tekeredve alkotja a kromatint. A kromatin szerkezetének lazítása (pl. hiszton acetilációval) hozzáférhetővé teszi a géneket a transzkripciós gépezet számára, míg a tömörítés (pl. DNS metilációval) elzárja azokat.
- RNS-feldolgozási szintű szabályozás (poszt-transzkripciós): Eukariótákban a pre-mRNS módosításai is szabályozhatók. A legfontosabb ilyen mechanizmus az alternatív splicing, amelyről már esett szó. Egy génről különböző splicing mintázatokkal több különböző mRNS és így több különböző fehérje is létrejöhet, növelve a fehérjék sokféleségét.
- Transzlációs szintű szabályozás: A sejt szabályozhatja azt is, hogy egy már elkészült mRNS molekula lefordítódik-e fehérjévé, és milyen hatékonysággal.
- mRNS stabilitás: Az mRNS molekulák élettartama változó. Az mRNS lebontását szabályozó mechanizmusok befolyásolják, hogy mennyi ideig áll rendelkezésre az mRNS a transzlációhoz.
- mikroRNS-ek (miRNA): Ezek a kis, nem kódoló RNS molekulák komplementer módon kötődnek specifikus mRNS-ekhez, és gátolják azok transzlációját, vagy lebontásra ítélik őket. Ez egy igen elterjedt és finom szabályozási mechanizmus.
- Iniciáció szabályozása: A transzláció iniciációs faktorainak aktivitásának szabályozása befolyásolhatja a fehérjeszintézis általános sebességét.
- Poszt-transzlációs szintű szabályozás: Még a kész fehérjék aktivitása is szabályozható, általában a poszt-transzlációs módosítások révén.
- Aktiváció/inaktiváció: A foszforiláció, glikoziláció és más kovalens módosítások ki- vagy bekapcsolhatják a fehérjék aktivitását, vagy megváltoztathatják funkciójukat.
- Fehérje lebontás: Az ubikvitin-proteaszóma rendszer szabályozza a fehérjék élettartamát. A fehérjék célzott lebontása kulcsfontosságú a sejtciklus szabályozásában, a jelátvitelben és a minőség-ellenőrzésben.
- Célba juttatás: A fehérjék megfelelő lokalizációja is egyfajta szabályozás, hiszen egy enzim csak akkor tud működni, ha a megfelelő kompartmentben van.
Példaként említhető a lac operon prokariótákban, amely a laktóz metabolizmusához szükséges géneket szabályozza. Laktóz hiányában egy represszor fehérje kötődik az operátorhoz, gátolva a transzkripciót. Laktóz jelenlétében a laktóz kötődik a represszorhoz, megváltoztatva annak konformációját, így az leválik a DNS-ről, és a gének expresszálódhatnak. Ez egy klasszikus példa a transzkripciós szintű szabályozásra.
Eukariótákban a génexpresszió szabályozása sokkal komplexebb, és alapvető a fejlődésbiológiában. A magzati fejlődés során a sejtek differenciálódnak, és különböző géneket fejeznek ki, hogy különböző sejttípusokká váljanak. A hormonális szabályozás is nagyrészt a génexpresszió befolyásolásán keresztül valósul meg, például az ösztrogén vagy a tesztoszteron számos gén transzkripcióját aktiválja a célsejtekben.
„A génexpresszió szabályozása a sejt intelligenciájának megnyilvánulása, mely lehetővé teszi, hogy minden pillanatban a legoptimálisabb módon reagáljon a belső és külső kihívásokra.”
A génexpresszió szabályozásának megértése alapvető fontosságú a biológia és az orvostudomány számos területén, a betegségek molekuláris mechanizmusainak felderítésétől kezdve a génterápiás beavatkozások fejlesztéséig.
A fehérjeszintézis zavarai és következményei
A fehérjeszintézis rendkívüli pontossága ellenére is előfordulhatnak hibák, amelyek súlyos következményekkel járhatnak a sejt és az egész szervezet számára. Bármelyik lépésben bekövetkező zavar, legyen az a transzkripció, a transzláció, a fehérjehajtogatás vagy a poszt-transzlációs módosítások szintjén, a fehérjék hibás működéséhez vagy hiányához vezethet, ami számos betegség alapját képezi.
A leggyakoribb problémák forrása a mutáció, azaz a DNS bázissorrendjének megváltozása. Mivel a DNS a fehérjeszintézis kiindulópontja, a DNS-ben bekövetkező bármilyen változás befolyásolhatja az mRNS szekvenciáját, ami végső soron egy hibás aminosav sorrendet eredményezhet.
- Pontmutációk: Egyetlen bázispár megváltozása a DNS-ben.
- Helyettesítő mutáció (missense mutation): Egyetlen aminosav megváltozását okozza a fehérjében. Példa erre a sarlósejtes anémia, ahol a hemoglobin béta-láncában egyetlen glutamin aminosavat valin helyettesít, ami a vörösvértestek sarló alakúvá válásához vezet, oxigénhiányos állapotban.
- Nonsense mutáció: Egy aminosavat kódoló kodon stop kodonná változik. Ez egy rövidebb, csonka fehérjét eredményez, amely gyakran teljesen funkcióképtelen.
- Synonymous (silent) mutation: A mutáció egy olyan kodonra változtatja a bázispárt, amely ugyanazt az aminosavat kódolja a genetikai kód degenerációja miatt. Ez nem okoz változást a fehérjében.
- Frameshift mutációk: Bázisok beékelődése (inzertáció) vagy kiesése (deléció), amelyek nem három többszörösei. Ezek a mutációk eltolják az olvasási keretet, ami a mutáció pontjától kezdve az összes további kodon megváltozását eredményezi, és általában teljesen funkcióképtelen fehérjét hoz létre.
A fehérjehajtogatás zavarai is komoly betegségeket okozhatnak. Ha egy fehérje nem veszi fel a megfelelő térszerkezetet, akkor általában funkcióképtelen, és gyakran hajlamos az aggregációra, káros lerakódásokat képezve a sejtekben és szövetekben.
- Cisztás fibrózis: Ezt a súlyos örökletes betegséget a CFTR (Cystic Fibrosis Transmembrane Conductance Regulator) fehérje hibás hajtogatása okozza. A leggyakoribb mutáció (ΔF508) egy fenilalanin aminosav hiányát eredményezi, ami a CFTR fehérje helytelen hajtogatásához és lebontásához vezet, mielőtt elérné a sejtfelszínt, ahol kloridion-csatornaként működne. Ez a klorid transzport zavarát okozza, ami sűrű nyák termelődéséhez vezet a tüdőben, hasnyálmirigyben és más szervekben.
- Alzheimer-kór és Parkinson-kór: Ezek a neurodegeneratív betegségek a tévesen hajtogatott fehérjék (pl. béta-amiloid, tau fehérje az Alzheimer-kórban; alfa-szinuklein a Parkinson-kórban) aggregációjával és lerakódásával járnak az agyban, ami idegsejtpusztuláshoz vezet.
- Prion betegségek: Rendkívül ritka és halálos betegségek (pl. Creutzfeldt-Jakob-kór), amelyeket egy normális prionfehérje (PrPC) tévesen hajtogatott, fertőző formájának (PrPSc) terjedése okoz. A PrPSc képes a normális PrPC fehérjéket is tévesen hajtogatott formává alakítani, láncreakciót indítva el, ami spongiform (szivacsos) agykárosodáshoz vezet.
A poszt-transzlációs módosítások hibái is kihatással lehetnek. Például a glikoziláció zavarai veleszületett glikozilációs rendellenességeket (CDG – Congenital Disorders of Glycosylation) okozhatnak, amelyek számos szervrendszert érinthetnek, a fejlődési rendellenességektől kezdve az idegrendszeri problémákig.
A fehérjeszintézis zavarainak megértése kulcsfontosságú a gyógyszerfejlesztés szempontjából is. Számos antibiotikum célzottan gátolja a baktériumok fehérjeszintézisét, anélkül, hogy károsítaná az emberi sejteket. Például a tetraciklinek gátolják a tRNS kötődését az A-helyhez, az eritromicin gátolja a transzlokációt, a kloramfenikol pedig a peptidkötés képződését. Mivel a prokarióta és eukarióta riboszómák szerkezete és működése között vannak különbségek, ezek az antibiotikumok szelektíven hatnak a baktériumokra.
„Minden hibásan hajtogatott fehérje egy elvesztett funkciót jelent, egy potenciális betegség magját, mely rávilágít a molekuláris precizitás életbe vágó fontosságára.”
A biotechnológiai alkalmazások terén a fehérjeszintézis szabályozásának ismerete lehetővé teszi, hogy baktériumokat vagy élesztőket használjunk emberi fehérjék, például inzulin vagy növekedési hormon nagy mennyiségben történő előállítására. Ez forradalmasította a gyógyszergyártást és számos betegség kezelését. A rekombináns DNS technológiák segítségével be tudjuk juttatni egy génünket egy baktérium sejtbe, ami aztán elkezdi termelni a kívánt emberi fehérjét.
A vakcinák előállítása is gyakran magában foglalja a fehérjeszintézist. Sok vakcina inaktivált vírust vagy annak egy részét, például egy specifikus fehérjéjét (antigénjét) tartalmazza. Ezeket a fehérjéket gyakran rekombináns technológiával, sejtkultúrákban állítják elő, hogy biztonságosan és hatékonyan stimulálják az immunválaszt.
A fehérjeszintézis hibáinak és zavarainak tanulmányozása nemcsak a betegségek megértéséhez vezet, hanem új terápiás stratégiák kidolgozásához is, a hibás fehérjék korrigálásától kezdve a lebontásuk elősegítéséig vagy a hiányzó funkció pótlásáig.
A fehérjeszintézis kutatásának jövője és új távlatok
A fehérjeszintézis alapjainak felfedezése, amelyért több Nobel-díjat is odaítéltek, alapjaiban változtatta meg a biológiáról alkotott képünket. Azonban a kutatás ezen a területen korántsem áll meg, sőt, folyamatosan új és izgalmas távlatokat nyit meg. A modern technológiák és a multidiszciplináris megközelítések lehetővé teszik, hogy egyre mélyebben megértsük ezt a komplex folyamatot, és új alkalmazási lehetőségeket fedezzünk fel az orvostudományban, a biotechnológiában és az iparban.
Az egyik legforradalmibb terület a szintetikus biológia és a mesterséges fehérjeszintézis. A tudósok ma már képesek mesterséges géneket tervezni és szintetizálni, majd ezeket bejuttatni sejtekbe, hogy olyan fehérjéket állítsanak elő, amelyek a természetben nem léteznek, vagy módosított funkciókkal rendelkeznek. Ez megnyitja az utat új enzimek, gyógyszerek és biomolekulák létrehozása előtt, amelyek specifikus ipari vagy orvosi célokra használhatók fel. A teljesen mesterséges genomok tervezése és beültetése élő sejtekbe már nem tudományos-fantasztikum, hanem valóság, amely alapjaiban változtathatja meg a gyógyszergyártást és a bioüzemanyag-termelést.
A génszerkesztés, különösen a CRISPR/Cas9 technológia megjelenése, új korszakot nyitott a fehérjeszintézis kutatásában és manipulációjában. A CRISPR lehetővé teszi a tudósok számára, hogy rendkívüli pontossággal módosítsák a DNS-t, akár egyetlen bázispárt is kicserélve vagy beillesztve. Ez nemcsak a betegségeket okozó mutációk kijavítására ad reményt a génterápiában, hanem lehetővé teszi a fehérjék tervezését is. A tudósok módosíthatják a géneket, hogy olyan fehérjéket hozzanak létre, amelyek megváltozott aktivitással, stabilitással vagy kölcsönhatási tulajdonságokkal rendelkeznek, ezzel feltárva a fehérje funkciójának és szerkezetének összefüggéseit.
A fehérjetervezés egyre inkább kulcsfontosságúvá válik a gyógyszerfejlesztésben. A hagyományos gyógyszerek gyakran kis molekulák, amelyek valamilyen fehérjéhez kötődve fejtik ki hatásukat. A fehérjetervezés azonban lehetővé teszi, hogy olyan teljesen új fehérjéket hozzanak létre, amelyek specifikusan kötődnek betegségeket okozó molekulákhoz, vagy enzimatikus aktivitással rendelkeznek, és célzottan avatkoznak be a kóros folyamatokba. Például, tervezett antitestek (monoklonális antitestek) már ma is széles körben alkalmazottak rák és autoimmun betegségek kezelésében, de a jövőben még specifikusabb és hatékonyabb fehérjék tervezésére számíthatunk.
A fehérje-fehérje kölcsönhatások vizsgálata is egyre hangsúlyosabbá válik. A sejtekben a fehérjék ritkán működnek izoláltan. Ehelyett komplex hálózatokban kommunikálnak és működnek együtt. A kölcsönhatások feltérképezése elengedhetetlen a sejtműködés átfogó megértéséhez és betegségek, például a rák vagy a fertőző betegségek molekuláris mechanizmusainak azonosításához. Új technológiák, mint a tömegspektrometria és a képalkotó eljárások, lehetővé teszik ezen komplex hálózatok dinamikus vizsgálatát.
A proteomika, a proteom (egy adott sejt vagy szervezet összes fehérjéjének összessége) tanulmányozása, a genomika (a gének tanulmányozása) után a következő nagy lépés a biológiai kutatásban. Míg a genom megmondja, hogy milyen fehérjékre van potenciálisan szükség, addig a proteomika feltárja, hogy mely fehérjék vannak valóban jelen, milyen mennyiségben, hol helyezkednek el, milyen módosításokon estek át, és hogyan lépnek kölcsönhatásba. Ez kulcsfontosságú a betegségek biomarkereinek azonosításában és a személyre szabott orvoslás fejlesztésében, ahol a kezeléseket az egyén egyedi fehérje profiljához igazítják.
„A fehérjeszintézis kutatásának jövője nem csupán a megértés mélyítéséről szól, hanem arról is, hogy miként tudjuk ezt a molekuláris gépezetet a saját céljainkra fordítani, forradalmasítva az orvostudományt és a biotechnológiát.”
Végül, a mesterséges intelligencia és a gépi tanulás egyre nagyobb szerepet játszik a fehérjeszintézis kutatásában. Ezek az eszközök segíthetnek előre jelezni a fehérjék térszerkezetét az aminosav sorrendjük alapján (pl. AlphaFold), optimalizálni a génszerkesztési stratégiákat, és azonosítani a gyógyszerfejlesztés új célpontjait. A fehérjeszintézis megértése és manipulálása az élet alapvető folyamatainak megértését és az emberi egészség javítását ígéri a jövőben.







