
A fotoszintézis, a földi élet alapja, a napfény energiáját használja fel a szén-dioxid és a víz cukrokká történő átalakítására. Ez a rendkívül komplex biokémiai folyamat a növényvilágban számos variációban valósul meg, alkalmazkodva a legkülönfélébb környezeti kihívásokhoz. Míg a növények többsége a klasszikus C3-as fotoszintézist folytatja, addig egy kisebb, de annál sikeresebb csoport a C4 fotoszintézis speciális útját választotta. Ez a kivételes mechanizmus lehetővé teszi számukra, hogy a forró, száraz és intenzív napfényes környezetben is kiemelkedő hatékonysággal működjenek, jelentős agronómiai és ökológiai előnyökre téve szert.
A C3 fotoszintézis korlátai és a fotolégzés jelensége
A fotoszintézis legismertebb és legelterjedtebb formája a C3 fotoszintézis. Nevét onnan kapta, hogy a szén-dioxid elsődleges fixálása során egy három szénatomos vegyület, a 3-foszfoglicerát (3-PGA) keletkezik. Ennek a folyamatnak a kulcsenzime a ribulóz-1,5-biszfoszfát-karboxiláz/oxigenáz, vagy röviden Rubisco. A Rubisco egy rendkívül ősi enzim, amely évmilliárdokon keresztül kulcsszerepet játszott az élet fejlődésében, de van egy jelentős hátránya, ami bizonyos körülmények között korlátozza a fotoszintézis hatékonyságát.
A Rubisco enzim, ahogy a neve is mutatja, nemcsak szén-dioxidot képes megkötni (karboxiláz aktivitás), hanem oxigént is (oxigenáz aktivitás). Amikor az enzim oxigénnel reagál, beindul a fotolégzésnek nevezett folyamat. Ez a reakció szén-dioxidot szabadít fel és energiát fogyaszt, anélkül, hogy cukrot termelne. Gyakorlatilag egy kontraproduktív folyamat, amely csökkenti a nettó fotoszintézis sebességét. A fotolégzés különösen intenzívvé válik magas hőmérsékleten, intenzív fényviszonyok között és alacsony intracelluláris szén-dioxid koncentrációnál, amikor a sztómák záródnak a vízpazarlás elkerülése érdekében.
A forró és száraz éghajlaton a C3 növényeknek komoly kihívásokkal kell szembenézniük. A vízpazarlás minimalizálása érdekében a sztómáikat gyakran zárva tartják, ami csökkenti a szén-dioxid beáramlását a levelekbe. Ez a belső CO2-koncentráció csökkenésével jár, miközben az oxigén koncentrációja a fotoszintézis során termelt O2 felhalmozódása miatt nő. Ilyen körülmények között a Rubisco egyre inkább az oxigenáz aktivitás felé hajlik, ami drámai mértékben növeli a fotolégzést és csökkenti a fotoszintézis nettó hatékonyságát. A növények szó szerint elégetik a már megtermelt cukrok egy részét, miközben CO2-t veszítenek.
„A fotolégzés, bár korábban haszontalannak vagy károsnak tartották, ma már egy komplex biokémiai útként ismert, amely szerepet játszhat a nitrogén anyagcserében és a stresszválaszban, de a szénfixáció hatékonyságát tekintve kétségkívül hátrányos.”
A C4 evolúciós válasz: egy hatékonyabb szénfixációs stratégia
A C4 fotoszintézis egy lenyűgöző evolúciós adaptáció, amely kiküszöböli a Rubisco oxigenáz aktivitásából eredő problémákat. Ez a speciális útvonal lehetővé teszi a növények számára, hogy még magas hőmérsékleten, intenzív fényviszonyok között és korlátozott vízellátás mellett is rendkívül hatékonyan kössék meg a szén-dioxidot. A C4-es növények a földi vegetáció mindössze mintegy 3%-át teszik ki, mégis a globális primer produkció körülbelül 20-30%-áért felelősek, ami jól mutatja kivételes teljesítményüket.
A C4 mechanizmus lényegében egy CO2-koncentráló pumpa, amely egy speciális anatómiai elrendezéssel párosulva biztosítja, hogy a Rubisco enzim mindig magas szén-dioxid koncentrációjú környezetben működhessen. Ez a stratégia minimalizálja a fotolégzést, és maximalizálja a szénfixáció sebességét, különösen kedvezőtlen körülmények között. Ennek köszönhetően a C4 növények jellemzően gyorsabban növekednek, és magasabb biomassza hozammal rendelkeznek, mint C3-as társaik hasonló környezetben.
A C4 fotoszintézis nem egyetlen evolúciós esemény eredménye, hanem számos alkalommal, függetlenül alakult ki különböző növénycsaládokban. Ez a konvergens evolúció ékes példája, ahol hasonló ökológiai nyomás hatására hasonló adaptációk jönnek létre. Jelenleg több mint 60 növénycsaládban azonosítottak C4-es fajokat, ami aláhúzza ennek a stratégiai útvonalnak a sikerességét és alkalmazkodóképességét.
A C4 fotoszintézis biokémiai útvonala: kétlépcsős szénfixáció
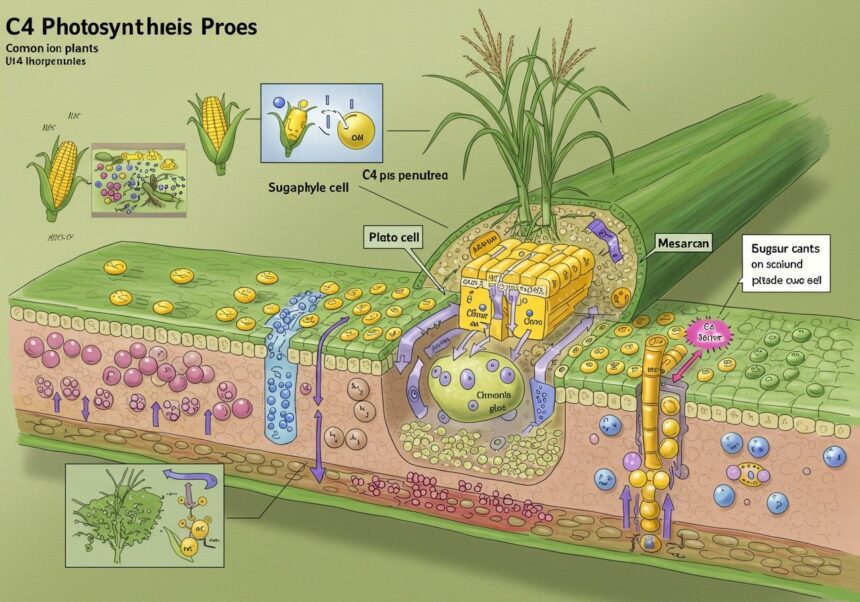
A C4 fotoszintézis a szén-dioxid fixálását térben és időben elkülönítve végzi, két fő sejttípus között elosztva a feladatokat: a mezofil sejtek és a kötegsejtek (bundle sheath cells) között. Ez a kétlépcsős folyamat biztosítja a Rubisco számára az ideális működési környezetet.
Az első lépés: CO2 fixáció a mezofil sejtekben
A légköri szén-dioxid először a levél külső részén elhelyezkedő mezofil sejtekbe diffundál. Itt találkozik a foszfoenolpiruvát-karboxiláz (PEP karboxiláz) enzimmel. Ez az enzim a foszfoenolpiruvátot (PEP) és a szén-dioxidot egy négy szénatomos vegyületté, az oxálacetáttá alakítja. A PEP karboxiláz kulcsfontosságú a C4 fotoszintézisben, mivel rendkívül nagy affinitással rendelkezik a szén-dioxid iránt, és nem reagál az oxigénnel. Ez azt jelenti, hogy még alacsony CO2-koncentrációnál is hatékonyan képes megkötni a szén-dioxidot, anélkül, hogy fotolégzés indulna be.
Az oxálacetát ezután gyorsan átalakul egy másik négy szénatomos vegyületté, jellemzően maláttá vagy aszpartáttá. A pontos vegyület attól függ, hogy a C4 útvonal melyik alaptípusáról van szó (NAD-ME, NADP-ME vagy PCK típus). Ezek a négy szénatomos savak szolgálnak a szén-dioxid „szállító molekuláiként”. Fontos megjegyezni, hogy ez a kezdeti fixálás nem igényel ATP-t a szén-dioxid megkötéséhez, de a PEP regenerálásához később szükség lesz rá.
A második lépés: CO2 koncentrálás és a Calvin-ciklus a kötegsejtekben
A malát vagy aszpartát molekulák a mezofil sejtekből aktív transzporttal, plazmodezmákon keresztül jutnak el a belső, érnyalábok körüli kötegsejtekbe. Ezek a sejtek vastagabb falúak és gyakran kevesebb oxigénnel érintkeznek, mint a mezofil sejtek. A kötegsejtekben a négy szénatomos savak dekarboxileződnek, azaz a szén-dioxid felszabadul belőlük. Ezzel egy időben egy három szénatomos vegyület, a piruvát keletkezik.
A felszabadult szén-dioxid koncentrációja a kötegsejtekben rendkívül magasra emelkedik, akár 10-12-szeresére is meghaladhatja a légköri CO2-koncentrációt. Ez a magas CO2-szint biztosítja az ideális környezetet a Rubisco enzim számára, amely itt, a kötegsejtek kloroplasztjaiban végzi a szén-dioxid végleges fixálását a Calvin-ciklusba. A Rubisco így szinte kizárólag a karboxiláz aktivitását tudja kifejteni, minimalizálva a fotolégzést.
A piruvát ezután visszakerül a mezofil sejtekbe, ahol ATP felhasználásával regenerálódik foszfoenolpiruváttá (PEP), készen arra, hogy újabb szén-dioxid molekulát kössön meg. Ez a regenerációs lépés biztosítja a C4 ciklus folyamatos működését, de egyúttal energiaigényes is, ami a C4 fotoszintézis egyik „költsége”.
A C4 fotoszintézis főbb típusai a dekarboxiláció enzime alapján
Bár az alapelv hasonló, a dekarboxiláció módja és az azt végző enzim alapján három fő C4 típus különböztethető meg:
- NADP-malát dehidrogenáz típus (NADP-ME): Ez a leggyakoribb típus, melyben a malát dekarboxilációját a NADP-függő malát dehidrogenáz enzim végzi. Példák: kukorica, cukornád.
- NAD-malát dehidrogenáz típus (NAD-ME): Ebben a típusban a malát dekarboxilációját a NAD-függő malát dehidrogenáz végzi. Példák: amaránt, köles.
- PEP karboxikináz típus (PCK): Itt a dekarboxilációt a PEP karboxikináz katalizálja, közvetlenül piruvátot és CO2-t hozva létre a PEP-ből. Példák: Guinea fű (Panicum maximum).
Ezek a típusok kisebb biokémiai különbségeket mutatnak, de mindegyikük ugyanazt a célt szolgálja: a szén-dioxid koncentrálását a Rubisco működési helyén, a kötegsejtekben.
„A C4 fotoszintézis a növények egyik leginnovatívabb adaptációja, amely egy komplex biokémiai „szén-dioxid pumpát” és egy speciális anatómiai elrendezést ötvözve teszi lehetővé a túlélést és a prosperálást a legmostohább körülmények között is.”
A Kranz anatómia: a C4 fotoszintézis strukturális alapja
A C4 fotoszintézis biokémiai hatékonysága elválaszthatatlanul összefonódik egy jellegzetes levélanatómiai struktúrával, amelyet Kranz anatómiának neveznek (a „Kranz” németül koszorút jelent). Ez az anatómiai elrendezés teszi lehetővé a kétlépcsős szénfixációs folyamat térbeli elkülönítését és a hatékony anyagszállítást a sejtek között.
A Kranz anatómia legfőbb jellemzője az érnyalábokat (vezetéknyalábokat) körülvevő, sűrűn elhelyezkedő és gyakran megnagyobbodott kötegsejtek egy vagy több rétege. Ezek a sejtek koncentrikusan, koszorúszerűen rendeződnek el az érnyalábok körül. Kívülről ezeket a kötegsejteket veszik körül a mezofil sejtek, amelyek lazább elrendezésűek és nagyobb felületen érintkeznek a levegővel.
A Kranz anatómia kulcsfontosságú jellemzői:
- Két sejttípus specializációja: A mezofil sejtek és a kötegsejtek morfológiailag és funkcionálisan is eltérnek. A mezofil sejtek felelősek a kezdeti CO2 fixációért (PEP karboxiláz), míg a kötegsejtekben történik a dekarboxiláció és a Calvin-ciklus (Rubisco).
- Kloroplasztok elhelyezkedése és típusa: A mezofil sejtek kloroplasztjai általában fejlett granumokkal rendelkeznek, és az elektron transzport lánc fényreakcióit végzik. Ezzel szemben a kötegsejtek kloroplasztjai gyakran nagyobbak, kevesebb vagy hiányzó granummal (agranális kloroplasztok) rendelkeznek, és specializálódtak a Calvin-ciklusra.
- Vastag sejtfal a kötegsejtek körül: A kötegsejtek vastagabb sejtfalakkal rendelkeznek, és gyakran szuberin tartalmú lamellákkal vannak szigetelve. Ez a struktúra csökkenti a gázok (különösen a CO2) kiszökését a kötegsejtekből a mezofil sejtekbe vagy a levél légtereibe, ezzel fenntartva a magas CO2-koncentrációt a Rubisco számára.
- Szoros kapcsolat a mezofil és kötegsejtek között: A két sejttípus között nagyszámú plazmodezma található, amelyek hatékony anyagszállítást biztosítanak (pl. malát vagy aszpartát transzportja a mezofil sejtekből a kötegsejtekbe, piruvát vissza).
- Rövid diffúziós utak: A szoros elrendezés miatt a gázok és metabolitok diffúziós távolsága minimális, ami hozzájárul a folyamat hatékonyságához.
A Kranz anatómia nem csak egy morfológiai érdekesség, hanem a C4 fotoszintézis működésének elengedhetetlen feltétele. Ez a speciális sejtelrendezés hozza létre azt a mikroklímát a kötegsejtekben, ahol a Rubisco enzim optimálisan, fotolégzés nélkül működhet.
A C4 fotoszintézis előnyei: miért olyan sikeres ez az út?
A C4 fotoszintézis számos jelentős előnnyel jár a növények számára, különösen bizonyos környezeti feltételek mellett. Ezek az előnyök magyarázzák a C4 növények dominanciáját a trópusi és szubtrópusi régiókban, valamint a mezőgazdaságban betöltött fontos szerepüket.
1. Magasabb fotoszintetikus hatékonyság
A C4 növények sokkal hatékonyabban képesek fotoszintetizálni magas hőmérsékleten és intenzív fényviszonyok között, mint a C3 növények. A CO2-koncentráló mechanizmusnak köszönhetően a Rubisco enzim folyamatosan telített állapotban van szén-dioxiddal, így maximális sebességgel dolgozhat. Ezáltal a C4 növények nettó fotoszintézis sebessége lényegesen magasabb lehet, ami gyorsabb növekedést és nagyobb biomassza termelést eredményez.
2. Csökkentett fotolégzés
Ez az egyik legfőbb előnye a C4 útvonalnak. A szén-dioxid koncentrálása a kötegsejtekben, ahol a Rubisco található, gyakorlatilag elnyomja az enzim oxigenáz aktivitását. Míg a C3 növények akár 20-50%-os energiaveszteséget is szenvedhetnek a fotolégzés miatt, addig a C4 növényeknél ez a veszteség minimális, vagy szinte teljesen hiányzik. Ez az energia megtakarítás közvetlenül hozzájárul a magasabb termelékenységhez.
3. Javított vízfogyasztási hatékonyság
A C4 növények rendkívül hatékonyan gazdálkodnak a vízzel. Mivel a PEP karboxiláz nagy affinitással rendelkezik a CO2 iránt, a C4 növényeknek nem kell olyan szélesre nyitniuk a sztómáikat, hogy elegendő szén-dioxidot vegyenek fel. A sztómák részleges zárva tartása csökkenti a transzspirációt (vízgőz leadását), miközben a fotoszintézis sebessége továbbra is magas marad. Ennek eredményeként a vízfelhasználási hatékonyság (WUE – water use efficiency), azaz a fotoszintetizált biomassza egységére jutó elpárologtatott víz mennyisége lényegesen jobb a C4 növényeknél, mint a C3 növényeknél. Ez az adaptáció kulcsfontosságú a száraz, aszályos régiókban.
4. Hatékonyabb nitrogénfelhasználás
A Rubisco enzim a növények egyik legbőségesebben előforduló fehérjéje, és jelentős mennyiségű nitrogént tartalmaz. Mivel a C4 növények Rubiscója sokkal hatékonyabban működik a magas CO2-koncentrációnak köszönhetően, kevesebb Rubisco enzimre van szükségük ugyanannyi szén-dioxid fixálásához, mint a C3 növényeknek. Ezáltal a C4 növények kevesebb nitrogént igényelnek a fotoszintetikus apparátus felépítéséhez, ami jelentős előny a nitrogénszegény talajokon. A nitrogénfelhasználási hatékonyságuk is magasabb.
5. Alkalmazkodás magas hőmérséklethez és intenzív fényhez
A C4 fotoszintézis nemcsak tolerálja a magas hőmérsékletet, hanem optimálisan működik ilyen körülmények között. A magas hőmérséklet növeli a Rubisco oxigenáz aktivitását, de a C4 mechanizmus ezt a problémát kiküszöböli. Emellett a magas fényintenzitás, amely a C3 növényeknél fotokárosodáshoz vezethet, a C4 növényeknél teljes mértékben kihasználásra kerül, mivel a fotoszintetikus apparátusuk képes a nagy mennyiségű fényenergia hatékony átalakítására.
6. Gyorsabb növekedés és magasabb terméshozam
A fent említett előnyök összessége gyorsabb növekedési rátát és magasabb biomassza hozamot eredményez. Ez különösen fontos a mezőgazdasági növények, például a kukorica és a cukornád esetében, amelyek a világ élelmiszer- és bioüzemanyag-ellátásában kulcsszerepet játszanak.
A C4 fotoszintézis hátrányai és kompromisszumai
Bár a C4 fotoszintézis számos előnnyel jár, nem minden körülmények között optimális. Vannak bizonyos hátrányai és kompromisszumai, amelyek megmagyarázzák, miért nem váltotta fel teljesen a C3 utat, és miért dominálnak még mindig a C3 növények a mérsékelt égövi és hidegebb klímákon.
- Magasabb energiaigény: A C4 útvonal működtetése energiaigényesebb. A PEP regenerálásához a piruvátból ATP-re van szükség, és a metabolitok transzportja is energiafelhasználással jár. Becslések szerint a C4 útvonal további 2 ATP ekvivalens energiát igényel minden egyes fixált CO2 molekulánként, szemben a C3 útvonallal. Ez a többlet energiafelhasználás alacsony fényintenzitás vagy alacsony hőmérséklet mellett hátrányossá válhat.
- Alacsonyabb hatékonyság hideg és alacsony fényviszonyok között: A C4 enzimek, különösen a PEP karboxiláz, optimálisan magasabb hőmérsékleten működnek. Alacsony hőmérsékleten a C4 mechanizmus kevésbé hatékony, és a C3 növények jobban teljesíthetnek. Hasonlóképpen, alacsony fényviszonyok mellett a C4 útvonal extra energiaigénye miatt a C3 növények gazdaságosabban működnek.
- Komplexebb anatómia és biokémia: A Kranz anatómia és a kétlépcsős biokémiai útvonal kialakítása és fenntartása komplexebb genetikai és fiziológiai apparátust igényel. Ez a bonyolultság korlátozhatja a C4 növények elterjedését bizonyos ökológiai fülkékben.
- Kevesebb nitrogén hasznosítás alacsony nitrogénszinten: Bár a C4 növények nitrogénfelhasználási hatékonysága magasabb, ez elsősorban a fotoszintetikus apparátusra vonatkozik. Extrém nitrogénhiányos talajokon a C4 mechanizmus fenntartásához szükséges enzimek és struktúrák előállítása mégis kihívást jelenthet.
A C4 növénytípusok sokfélesége és ökológiai jelentőségük
A C4 fotoszintézis, ahogy már említettük, több mint 60 növénycsaládban fejlődött ki függetlenül, ami a konvergens evolúció egyik legszebb példája. Ez a sokféleség azt jelenti, hogy a C4 növények rendkívül változatosak morfológiailag és ökológiailag is, a fűféléktől a pozsgásokig.
Főbb C4 növénycsaládok és példáik:
- Poaceae (Pázsitfűfélék): Ez a család tartalmazza a legtöbb C4 fajt, beleértve a világ legfontosabb gabonaféléit.
- Kukorica (Zea mays): Az egyik legismertebb és gazdaságilag legfontosabb C4 növény. Magas terméshozama és alkalmazkodóképessége miatt alapvető élelmiszer és takarmány.
- Cukornád (Saccharum officinarum): A világ legnagyobb biomassza termelője, a cukor és a bioüzemanyag (etanol) fő forrása.
- Köles (Panicum spp., Setaria spp., Pennisetum spp.): Számos kölesfaj, mint például a gyöngyköles vagy a rókafarkú köles, C4-es. Fontosak a szárazabb régiókban.
- Sorghum (Sorghum bicolor): Gabonaféle, takarmánynövény és bioüzemanyag alapanyag, különösen ellenálló a szárazsággal szemben.
- Gyepfűfélék: Számos trópusi és szubtrópusi gyepfű, mint például a Bermuda fű (Cynodon dactylon) vagy a pázsitfűfélék, C4-esek, és dominálnak a legelőkön.
- Cyperaceae (Sásfélék): Számos sásfaj is alkalmazza a C4 utat, különösen a trópusi vizes élőhelyeken.
- Példák: Egyes Cyperus fajok.
- Amaranthaceae (Disznóparéjfélék): Ez a család számos C4-es gyomnövényt és kultúrnövényt is tartalmaz.
- Amaránt (Amaranthus spp.): Értékes gabonaféle és levélzöldség, szárazságtűrő és tápláló.
- Disznóparéj (Amaranthus retroflexus): Hírhedt gyomnövény, amely a C4 mechanizmusnak köszönhetően rendkívül versenyképes a kultúrnövényekkel szemben.
- Chenopodiaceae (Libatopfélék): Egyes fajok, különösen a sós, száraz élőhelyeken élők, C4-esek.
- Példák: Egyes Atriplex (laboda) fajok.
- Euphorbiaceae (Kutyatejfélék): Néhány faj ebben a családban is C4 fotoszintézist mutat.
- Példák: Egyes Euphorbia fajok.
Ökológiai jelentőség:
A C4 növények dominálnak a trópusi és szubtrópusi füves pusztákon, szavannákon és félsivatagos területeken, ahol a magas hőmérséklet, az intenzív napsugárzás és a korlátozott vízellátás a jellemző. A C4 fotoszintézis lehetővé teszi számukra, hogy ezeken a kihívásokkal teli élőhelyeken is sikeresen versenyezzenek a C3 növényekkel, sőt, gyakran felülmúlják azokat. A globális felmelegedés és a CO2-szint emelkedése kapcsán a C4 növények ökológiai szerepe és elterjedése is a kutatások fókuszában áll.
A C4 növények szerepe a mezőgazdaságban és az élelmiszerbiztonságban

A C4 növények kritikus szerepet játszanak a globális élelmiszer- és takarmánytermelésben, valamint a bioüzemanyag-iparban. A kiemelkedő termelékenységük és stressztűrő képességük miatt számos alapvető kultúrnövény tartozik ebbe a csoportba.
Főbb C4 kultúrnövények:
- Kukorica (Zea mays): A világ egyik legfontosabb gabonaféléje, alapvető élelmiszer, takarmány és ipari alapanyag. Magas biomassza termelése és széles körű alkalmazkodóképessége teszi rendkívül értékessé.
- Cukornád (Saccharum officinarum): A cukorgyártás elsődleges alapanyaga, emellett jelentős forrása a bioetanolnak, különösen Brazíliában. A legnagyobb fotoszintetikus hatékonyságú növények közé tartozik.
- Sorghum (Sorghum bicolor): Fontos gabonaféle az aszályos régiókban, ahol a kukorica nem termeszthető gazdaságosan. Takarmányként, élelmiszerként és bioüzemanyagként is használják.
- Kölesfélék: Számos kölesfaj (pl. gyöngyköles, ujjasköles) alapvető élelmiszer Afrikában és Ázsia szárazabb részein, magas tápértékük és szárazságtűrő képességük miatt.
C4 gyomnövények:
A C4-es gyomnövények, mint például a disznóparéj (Amaranthus spp.), a kakaslábfű (Echinochloa crus-galli) vagy a muhar (Setaria spp.), komoly kihívást jelentenek a mezőgazdaságban. A C4 fotoszintézis adta versenyelőnyük (gyors növekedés, hatékony víz- és nitrogénhasználat) miatt rendkívül nehéz őket kiirtani, és jelentős termésveszteséget okozhatnak. Ezért a gyomirtási stratégiák kidolgozásában figyelembe kell venni a C4 mechanizmus sajátosságait.
A C4 rizs projekt:
Az élelmezésbiztonság növelése érdekében a tudósok egy ambiciózus projekten dolgoznak: a C4 rizs kifejlesztésén. A rizs (Oryza sativa) egy C3 növény, amely a világ lakosságának jelentős részének alapvető élelmiszere. Azonban a C3 fotoszintézis korlátai miatt a rizs terméshozama nem mindig optimális, különösen a melegebb éghajlatokon. A C4 rizs projekt célja, hogy genetikai módosítással bevezesse a C4 fotoszintézist a rizsbe, ezzel akár 50%-kal növelve a terméshozamot, javítva a nitrogén- és vízfelhasználási hatékonyságot. Ez hatalmas áttörést jelenthet a globális élelmiszerbiztonságban, különösen a klímaváltozás hatásainak tükrében.
A C4 fotoszintézis evolúciója: egy sokszorosan kialakult adaptáció
A C4 fotoszintézis evolúciója az egyik legizgalmasabb és legintenzívebben kutatott terület a növénybiológiában. Ahogy már említettük, ez a komplex adaptáció nem egyszer, hanem legalább 62, de valószínűleg több mint 60-szor alakult ki függetlenül, különböző növénycsaládokban az elmúlt 30 millió évben. Ez a konvergens evolúció jelensége rávilágít arra, hogy a C4 útvonal milyen erős szelekciós előnnyel járt bizonyos környezeti feltételek mellett.
Az evolúciós hajtóerők:
A C4 fotoszintézis kialakulását valószínűleg több tényező együttesen vezérelte:
- Alacsony légköri CO2-szint: A földtörténeti korokban, különösen az oligocén és miocén időszakokban, a légköri CO2-szint jelentősen alacsonyabb volt, mint ma. Ez a Rubisco oxigenáz aktivitásának növekedését és a fotolégzés intenzitásának fokozódását eredményezte a C3 növényeknél, ami szelekciós nyomást gyakorolt egy hatékonyabb CO2-koncentráló mechanizmus kialakítására.
- Magas hőmérséklet és szárazság: A globális klíma felmelegedése és az ariditás növekedése a Rubisco oxigenáz aktivitását is fokozta. A C4 útvonal különösen hatékonyan működik magas hőmérsékleten és száraz körülmények között, ahol a C3 növények szenvednek a vízpazarlás és a fotolégzés miatt.
- Intenzív napfény: A magas fényintenzitás szintén hozzájárult a C4 evolúciójához, mivel a C4 növények képesek a nagy mennyiségű fényenergia hatékonyabb hasznosítására, anélkül, hogy fotokárosodást szenvednének.
Az evolúciós lépcsők:
A C4 útvonal kialakulása nem egy hirtelen esemény volt, hanem fokozatosan, több lépcsőben ment végbe. A kutatók úgy vélik, hogy az evolúció során a C3 növényekből először C3-C4 intermedierek alakultak ki. Ezek a növények már mutattak bizonyos C4-es jellegzetességeket, például a fotolégzés csökkenését, de még nem rendelkeztek teljesen kifejlett Kranz anatómiával vagy a teljes C4 biokémiai útvonallal. Ezek az intermedierek valószínűleg átmeneti formák voltak, amelyek előkészítették a terepet a teljes C4 mechanizmus kialakulásához.
Az evolúciós folyamat során a meglévő C3-as gének és enzimek funkciói módosultak, és új gének is megjelentek (pl. a PEP karboxiláz génjeinek duplikációja és diverzifikációja), amelyek lehetővé tették a két sejttípus közötti munkamegosztást és a szén-dioxid koncentrálását. Ez a komplex folyamat rávilágít a növények adaptációs képességére és arra, hogy a környezeti nyomás hogyan formálja a biológiai rendszereket.
C3, C4 és CAM fotoszintézis: összehasonlítás
A fotoszintézis három fő típusát különböztetjük meg a szén-dioxid fixálás mechanizmusa alapján: C3, C4 és CAM (Crassulacean Acid Metabolism). Mindegyik út a környezeti kihívásokra adott eltérő evolúciós válaszokat képviseli.
| Jellemző | C3 Fotoszintézis | C4 Fotoszintézis | CAM Fotoszintézis |
|---|---|---|---|
| Első fixált termék | 3-foszfoglicerát (3 szénatom) | Oxálacetát (4 szénatom) | Oxálacetát (4 szénatom) |
| Primer CO2 fixáló enzim | Rubisco | PEP karboxiláz (mezofil sejtekben) | PEP karboxiláz (éjszaka) |
| Szénfixáció térbeli/időbeli elkülönítése | Nincs elkülönítés | Térbeli elkülönítés (mezofil vs. kötegsejtek) | Időbeli elkülönítés (éjszaka vs. nappal) |
| Fotolégzés | Jelentős, különösen melegben | Minimális vagy nincs | Minimális vagy nincs |
| Vízfelhasználási hatékonyság (WUE) | Alacsony | Magas | Nagyon magas |
| Optimális hőmérséklet | 15-25 °C | 25-45 °C | Nagy hőmérséklet-ingadozást tolerál |
| Előfordulás | Mérsékelt égövi növények többsége (búza, rizs, szója) | Trópusi/szubtrópusi füvek, kukorica, cukornád, amaránt | Pozsgások, kaktuszok, ananász |
| Sztómák nyitása | Nappal | Nappal (részlegesen zárva) | Éjszaka |
A CAM fotoszintézis (Crassulacean Acid Metabolism) egy másik speciális út, amelyet elsősorban a pozsgás növények, kaktuszok és más szárazságtűrő fajok alkalmaznak. A CAM növények a C4-hez hasonlóan PEP karboxilázzal kötik meg a CO2-t, de ezt éjszaka teszik, amikor a sztómáik nyitva vannak, és a vízpazarlás minimális. A megkötött CO2-t malát formájában tárolják a vakuólumokban. Nappal, amikor a sztómák zárva vannak, a malát dekarboxileződik, és a felszabadult CO2 a Calvin-ciklusba lép. Ez az időbeli elkülönítés rendkívül magas vízfogyasztási hatékonyságot biztosít, lehetővé téve a túlélést extrém száraz és forró környezetben.
A C4 fotoszintézis jövője és a klímaváltozás
A klímaváltozás és a légköri CO2-szint emelkedése jelentős hatással van a növényvilágra és a mezőgazdaságra. A C4 fotoszintézis ezen változások tükrében kiemelt figyelmet kap.
A CO2-szint emelkedése általában kedvez a C3 növényeknek, mivel a Rubisco oxigenáz aktivitását csökkenti, így a fotolégzés mértéke is mérséklődik. Ezt nevezzük CO2-trágyázási hatásnak. A C4 növényeknél ez a hatás kevésbé kifejezett, mivel a Rubisco már alapból magas CO2-koncentrációban működik. Azonban a klímaváltozás során várhatóan emelkedő hőmérséklet és a szárazság növekedése a C4 növényeknek adhat előnyt, mivel ők jobban adaptálódtak ezekhez a körülményekhez.
A C4 növények kutatása és fejlesztése kulcsfontosságú lehet a jövő élelmezésbiztonságában. A már említett C4 rizs projekt mellett, a tudósok más C3 növényekbe is megpróbálják bejuttatni a C4 mechanizmust, vagy legalábbis annak egyes elemeit, hogy javítsák azok termelékenységét és stressztűrő képességét. Ez a géntechnológiai megközelítés óriási potenciált rejt magában a mezőgazdasági termelés optimalizálására a változó éghajlati viszonyok között. A C4 növények vizsgálata segít megérteni a fotoszintézis alapvető mechanizmusait, és új utakat nyit meg a növénytermesztés hatékonyságának növelésére.







