
Az élő rendszerek működésének alapja a molekuláris szintű kölcsönhatások precíz koreográfiája. Ezen bonyolult tánc egyik legkiemelkedőbb szereplője az enzim, amely katalizátorként felgyorsítja a biokémiai reakciókat, lehetővé téve az élet fenntartásához szükséges folyamatokat. Az enzimek specifikus működése, a szubsztrátokkal való rendkívül szelektív kötődése évtizedek óta foglalkoztatja a tudósokat, és számos modell született ennek magyarázatára.
A legkorábbi és talán legismertebb elmélet, az Emil Fischer által 1894-ben javasolt „zár és kulcs modell”, egyszerű és intuitív magyarázatot kínált az enzim-szubsztrát kölcsönhatásra. Eszerint az enzim aktív centruma és a szubsztrát tökéletesen illeszkednek egymáshoz, mint egy kulcs a zárba. Ez a modell kiválóan magyarázta az enzimek rendkívüli specifikusságát, azonban hamarosan kiderült, hogy számos megfigyelést nem képes maradéktalanul értelmezni, különösen az enzimek rugalmasságát és a reakciók dinamikáját illetően.
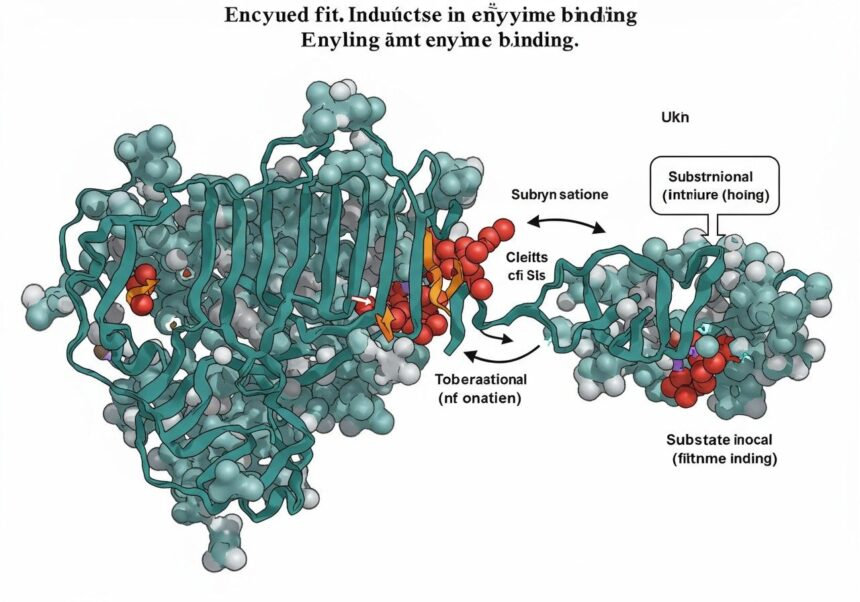
A 20. század közepén a tudomány egyre mélyebbre ásott a molekuláris struktúrák világába, és nyilvánvalóvá vált, hogy a fehérjék nem merev, statikus entitások, hanem dinamikus, mozgékony makromolekulák. E felismerés nyomán született meg Daniel Koshland Jr. által 1958-ban az indukált alkalmazkodási modell (induced fit model), amely forradalmasította az enzimkötődésről és -katalízisről alkotott képünket. Ez az elmélet nem csupán a szubsztrát és az enzim közötti kölcsönhatást írja le, hanem rávilágít a fehérjék inherent rugalmasságára és arra, hogy a kötődés során mindkét fél aktívan részt vesz az illeszkedés kialakításában.
Az indukált alkalmazkodási modell születése és alapelvei
A „zár és kulcs” modell korlátai egyre nyilvánvalóbbá váltak, ahogy a kutatók részletesebben vizsgálták az enzimek működését. A modell nem tudta kielégítően magyarázni például az enzimek allosztérikus szabályozását, ahol egy, az aktív centrumtól távoli helyen kötődő molekula befolyásolja az enzim aktivitását. Továbbá, bizonyos enzimek esetében megfigyelték, hogy a szubsztrát kötődése után az enzim szerkezetében jelentős változások mennek végbe, amelyek elengedhetetlenek a katalízishez.
Az indukált alkalmazkodási modell felismerte, hogy az enzim és a szubsztrát nem passzív partnerek, hanem aktívan formálják egymást a tökéletes illeszkedés eléréséért.
Daniel Koshland Jr., a neves biokémikus, felismerte, hogy az enzimek rugalmasabbak, mint azt korábban gondolták. Elmélete szerint a szubsztrát kötődése nem egy előre kialakított, statikus aktív centrumba történik, hanem maga a szubsztrát indukálja az enzim konformációjának megváltozását. Ez a változás alakítja ki az optimális aktív centrumot a katalízishez. Más szóval, az enzim alakja alkalmazkodik a szubsztráthoz, és a szubsztrát is enyhén módosulhat az enzimhez való kötődés során. Ez a kölcsönös alkalmazkodás maximalizálja a kötődés erejét és a katalitikus hatékonyságot.
Az indukált alkalmazkodási modell alapvető tézise, hogy az enzim aktív centruma nem egy merev, fix üreg, hanem egy dinamikus terület, amely képes megváltoztatni alakját és térbeli elrendeződését a szubsztrát jelenlétében. Ez a konformációs változás kulcsfontosságú a következő lépésekhez:
- Kötés optimalizálása: A kezdeti, gyenge szubsztrátkötődés kiváltja az enzim konformációs változását, ami megnöveli a kötődési affinitást és specifikusságot.
- Katalitikus csoportok elrendezése: A konformációs változás során az aktív centrumban lévő katalitikus aminosav oldalláncok optimális pozícióba kerülnek a reakció végrehajtásához.
- Víz kizárása: Gyakran a konformációs változás segít kizárni a vizet az aktív centrumból, ami megakadályozza a nem kívánt mellékreakciókat, és növeli a katalízis hatékonyságát.
- Átmeneti állapot stabilizálása: Az enzim alakváltozása hozzájárul az átmeneti állapot stabilizálásához, ami csökkenti az aktiválási energiát és felgyorsítja a reakciót.
Ez a dinamikus megközelítés sokkal rugalmasabb és valósághűbb képet fest az enzim-szubsztrát kölcsönhatásról, mint a korábbi statikus modellek. Az indukált alkalmazkodási modell nemcsak a katalízis mechanizmusát magyarázza meg jobban, hanem alapjául szolgál a modern gyógyszertervezésnek és az enzimfejlesztésnek is.
Az enzim szerkezetének dinamikája és a konformációs változások
Az enzimek, mint fehérjék, rendkívül komplex makromolekulák, amelyek háromdimenziós szerkezete határozza meg működésüket. Az indukált alkalmazkodási modell megértéséhez elengedhetetlen a fehérjék dinamikus természetének felismerése. A fehérjék molekulái nem merev struktúrák, hanem folyamatosan mozognak, rezegnek, forognak, és képesek nagyméretű konformációs változásokra. Ezek a mozgások alapvetőek a biológiai funkcióikhoz.
A fehérjék rugalmasságát számos tényező befolyásolja, többek között az aminosav-szekvencia, a másodlagos szerkezeti elemek (alfa-hélixek, béta-redők), a doménszerkezet és a diszulfidhidak jelenléte. Az enzimek gyakran több doménből állnak, amelyek viszonylagosan függetlenül mozoghatnak egymáshoz képest. Ez a doménmozgás (domain motion) kulcsfontosságú az indukált alkalmazkodás során.
Amikor egy szubsztrát közeledik az enzimhez, az első, gyenge kölcsönhatások (van der Waals erők, hidrogénkötések, ionos kötések) kiváltják az enzim szerkezetének finom átrendeződését. Ez a kezdeti változás további kölcsönhatásokat tesz lehetővé, ami erősíti a kötődést, és egyre nagyobb konformációs változásokat indukál. Ez a folyamat addig folytatódik, amíg az enzim és a szubsztrát el nem éri az optimális, átmeneti állapot-szerű konformációt.
A konformációs változások mértéke és típusa rendkívül változatos lehet. Néhány enzim esetében csak apró, lokalizált átrendeződések történnek az aktív centrum közvetlen közelében. Más esetekben, például a hexokináz enzimnél, az egész molekula jelentős, „szájzáró” mozgást végez, bezárva a szubsztrátot az aktív centrumba. Ez a makroszkopikus mozgás biztosítja, hogy a szubsztrát teljesen elszigetelődjön a környező víztől, és az aktív centrum katalitikus csoportjai optimálisan helyezkedjenek el.
Az enzim dinamikus természete, a folyamatos mozgás és a konformációs flexibilitás teszi lehetővé, hogy precízen reagáljon a szubsztrát jelenlétére és optimalizálja a katalitikus hatékonyságot.
A fehérje dinamika nem csupán az indukált alkalmazkodás mechanizmusát alapozza meg, hanem számos más biológiai folyamatban is kulcsszerepet játszik, mint például a jeltovábbításban, a fehérje-fehérje kölcsönhatásokban és a molekuláris gépek működésében. A konformációs változások vizsgálata modern biokémia és molekuláris biológia egyik legaktívabb kutatási területe.
Az indukált alkalmazkodás szerepe a katalízisben
Az enzimek fő feladata a kémiai reakciók sebességének növelése azáltal, hogy csökkentik az aktiválási energiát. Az indukált alkalmazkodási modell kulcsfontosságú annak megértésében, hogy az enzimek hogyan érik el ezt a figyelemre méltó hatékonyságot. A modell szerint az enzim nem csupán egy helyet biztosít a szubsztrát számára, hanem aktívan részt vesz a reakcióút átmeneti állapotának stabilizálásában.
Amikor a szubsztrát kötődik az enzimhez, az indukált konformációs változás következtében az aktív centrumban lévő katalitikus aminosav oldalláncok (pl. savas, bázikus, nukleofil vagy elektrofil csoportok) pontosan a megfelelő térbeli elrendeződésbe kerülnek. Ez az optimalizált pozíció lehetővé teszi, hogy ezek a csoportok hatékonyan kölcsönhatásba lépjenek a szubsztráttal, elősegítve a kovalens kötések képződését vagy felhasadását. Az enzim ilyenkor nemcsak „tartja” a szubsztrátot, hanem aktívan „segíti” a kémiai átalakulást.
Az átmeneti állapot stabilizálása az enzimkatalízis egyik legfontosabb elve. Az átmeneti állapot egy rendkívül instabil, rövid életű szerkezet, amely a reaktánsok és a termékek közötti úton található. Az enzimek úgy működnek, hogy sokkal erősebben és specifikusabban kötődnek az átmeneti állapothoz, mint a szubsztrátokhoz vagy a termékekhez. Az indukált alkalmazkodás ebben a folyamatban kulcsszerepet játszik, mivel az enzim konformációs változása pontosan olyan alakot vesz fel, amely komplementer az átmeneti állapot szerkezetével.
A kötődési energia, amelyet az enzim az átmeneti állapot kötésével nyer, felhasználódik az aktiválási energia csökkentésére. Ez a mechanizmus magyarázza az enzimek rendkívüli katalitikus hatékonyságát, amely akár milliószorosára is felgyorsíthatja a reakciókat. Az indukált alkalmazkodás tehát nem csupán a kötődés pontosságát biztosítja, hanem közvetlenül hozzájárul a katalitikus sebesség növeléséhez is.
Egy további fontos aspektus a víz kizárása az aktív centrumból. Számos biokémiai reakció víz jelenlétében is lejátszódhatna, de gyakran sokkal lassabban, és kevésbé specifikusan. Az indukált alkalmazkodás során az enzim „bezárja” a szubsztrátot, elzárva azt a környező víztől. Ez a dehidrált környezet kedvezőbbé teheti a reakciót, megakadályozhatja a nem kívánt hidrolízist, és növelheti a katalízis specificitását.
Az indukált alkalmazkodás nem csak a szubsztrátkötés pontosságát biztosítja, hanem aktívan hozzájárul az átmeneti állapot stabilizálásához, ami az enzimkatalízis hatékonyságának kulcsa.
Összefoglalva, az indukált alkalmazkodási modell szerint az enzim és a szubsztrát közötti dinamikus kölcsönhatás teszi lehetővé az optimális katalitikus környezet kialakítását, az átmeneti állapot hatékony stabilizálását, és ezáltal a reakciók rendkívüli felgyorsítását.
Példák az indukált alkalmazkodásra a természetben

Az indukált alkalmazkodási modell nem csupán egy elméleti konstrukció, hanem számos enzim működésében kísérletileg is igazolták. Ezek a példák segítenek megérteni, hogyan működik ez az elv a biológiai rendszerekben.
Hexokináz
Az egyik klasszikus és talán leggyakrabban idézett példa a hexokináz enzim, amely a glükóz foszforilációját katalizálja, egy ATP molekula felhasználásával. Ez az első lépés a glikolízisben. A hexokináz két doménből áll, amelyek egy „réssel” vannak elválasztva. A glükóz hiányában a rés nyitott, és az aktív centrum nem optimális a katalízishez.
Amikor a glükóz kötődik a hexokinázhoz, az enzim jelentős konformációs változáson megy keresztül: a két domén bezáródik egymásra, mint egy száj. Ez a „szájzáró” mozgás elszigeteli a glükózt a környező víztől, és optimális pozícióba hozza az ATP-t és a glükózt a foszfátcsoport átviteléhez. Ez a változás megakadályozza az ATP hidrolízisét is, ami egy fontos mellékreakció lenne víz jelenlétében. A hexokináz specifikusan a glükózra reagál ezzel a bezáródással; más cukrok, mint például a víz, nem váltják ki ezt a konformációs változást.
Alkohol-dehidrogenáz
Az alkohol-dehidrogenáz (ADH) egy másik kiváló példa, amely a NAD+ koenzim és az alkohol szubsztrát kötődésekor mutat indukált alkalmazkodást. Az ADH egy cinkiont tartalmazó enzim, amely az alkoholt aldehiddé oxidálja. A koenzim és a szubsztrát kötődésekor az enzim szerkezetében jelentős átrendeződések mennek végbe. Ezek a változások optimalizálják a katalitikus cinkion és a szubsztrát közötti távolságot és orientációt, elősegítve a hidrogénátvitelt.
Lizozim
A lizozim egy enzim, amely a baktériumok sejtfalának hidrolízisében játszik szerepet. Bár gyakran a „zár és kulcs” modell illusztrálására használják, mivel az aktív centruma jól illeszkedik a szubsztrátjához (egy poliszacharidhoz), valójában finom indukált alkalmazkodást is mutat. A szubsztrát kötődésekor az enzim aktív centrumában lévő bizonyos aminosav oldalláncok elmozdulnak, hogy jobban stabilizálják az átmeneti állapotot és elősegítsék a glikozidos kötés felhasadását. Ez a példa rámutat arra, hogy az indukált alkalmazkodás nem feltétlenül jelent drámai, nagyméretű konformációs változást; akár finom átrendeződések is elegendőek lehetnek.
DNS-polimeráz
A DNS-polimerázok, amelyek a DNS replikációjáért felelősek, szintén indukált alkalmazkodást mutatnak. Ezek az enzimek rendkívül pontosak a nukleotidok beépítésében, és ez a pontosság részben az indukált alkalmazkodásnak köszönhető. Amikor egy helyes nukleotid (komplementer a templát szállal) kötődik az aktív centrumba, az enzim „ujjai” bezáródnak, és egy zárt konformációt vesz fel. Ez a konformáció stabilizálja a helyes párosítást, és elősegíti a foszfodiészter kötés kialakulását. Ha egy hibás nukleotid próbál beépülni, az enzim nem veszi fel ezt a zárt konformációt, ami lehetővé teszi a hibás nukleotid eltávolítását, ezzel biztosítva a magas hűséget a replikáció során.
Ezek a példák jól illusztrálják, hogy az indukált alkalmazkodás egy univerzális mechanizmus, amely számos különböző enzim és biológiai folyamat alapját képezi, hozzájárulva azok specifikusságához és hatékonyságához.
Az indukált alkalmazkodás és az allosztérikus szabályozás
Az enzimek aktivitását az élő rendszerekben szigorúan szabályozzák, hogy a sejtek energia- és anyagcsere igényeinek megfelelően működjenek. Az egyik legfontosabb szabályozási mechanizmus az allosztérikus szabályozás, amely szorosan összefügg az indukált alkalmazkodási modellel. Az allosztérikus szabályozás során egy molekula (az allosztérikus effektor) egy, az aktív centrumtól eltérő helyen (az allosztérikus centrumban) kötődik az enzimhez, és ezzel megváltoztatja az enzim aktivitását.
Az allosztérikus effektor kötődése konformációs változást indukál az enzimben, amely az aktív centrumra is kiterjed. Ez a konformációs változás vagy növelheti (allosztérikus aktivátor), vagy csökkentheti (allosztérikus inhibitor) az enzim szubsztrátjához való affinitását, vagy a katalitikus sebességét. Az indukált alkalmazkodási modell kiválóan magyarázza ezt a „távoli” hatást, mivel feltételezi, hogy az enzim szerkezete rugalmas és képes az információt az egyik területről a másikra továbbítani.
Az allosztérikus enzimek gyakran több alegységből állnak, és az egyik alegységen bekövetkező konformációs változás befolyásolhatja a többi alegység szerkezetét és funkcióját. Ez az alegység-alegység közötti kommunikáció a kooperativitás alapja, amely például a hemoglobin oxigénkötésénél is megfigyelhető, bár a hemoglobin nem enzim. Enzimek esetében ez azt jelenti, hogy az allosztérikus effektor kötődése az egyik alegységhez megváltoztatja az összes alegység aktív centrumát, ezzel fokozva vagy gátolva az enzim teljes aktivitását.
Az allosztérikus szabályozás az indukált alkalmazkodás tökéletes példája, ahol egy távoli kötődés is képes az enzim teljes konformációját és aktivitását megváltoztatni, finomhangolva ezzel a biokémiai útvonalakat.
Az allosztérikus szabályozás rendkívül fontos az anyagcsere útvonalak finomhangolásában. Például, a glikolízis első lépését katalizáló foszfofruktokináz-1 (PFK-1) egy allosztérikus enzim, amelyet az ATP (egy inhibitor) és az AMP (egy aktivátor) szabályoz. Magas ATP-szint esetén az ATP allosztérikusan kötődik a PFK-1-hez, csökkentve annak aktivitását, és lassítva a glikolízist. Alacsony ATP-szint (magas AMP-szint) esetén az AMP aktiválja az enzimet, felgyorsítva a glikolízist az energia termelésére. Ez a mechanizmus biztosítja, hogy a glikolízis sebessége pontosan illeszkedjen a sejt energiaigényeihez.
Az allosztérikus szabályozás megértése kulcsfontosságú a gyógyszerfejlesztésben is. Az allosztérikus gyógyszerek, amelyek nem az aktív centrumba, hanem egy allosztérikus helyre kötődnek, gyakran specifikusabbak és kevesebb mellékhatással járnak, mivel kevésbé valószínű, hogy más, hasonló aktív centrummal rendelkező enzimeket is befolyásolnak.
Az indukált alkalmazkodás kísérleti bizonyítékai
Az indukált alkalmazkodási modell érvényességét számos kísérleti technika támasztja alá, amelyek lehetővé tették a kutatók számára, hogy molekuláris szinten figyeljék meg az enzimek konformációs változásait.
Röntgenkrisztallográfia
A röntgenkrisztallográfia az egyik legerősebb eszköz a fehérjék háromdimenziós szerkezetének meghatározására. Ennek a technikának köszönhetően a kutatók képesek voltak kristályosítani az enzimeket mind szubsztrát nélkül (apoenzim), mind szubsztráttal vagy szubsztrát analóggal kötött állapotban (holoenzim). A két szerkezet összehasonlítása egyértelműen kimutatta a szubsztrátkötés által indukált konformációs változásokat. A hexokináz „szájzáró” mozgását például részletesen dokumentálták röntgenkrisztallográfiás adatokkal.
Nukleáris mágneses rezonancia (NMR) spektroszkópia
Az NMR spektroszkópia lehetővé teszi a fehérjék dinamikájának és konformációs állapotainak vizsgálatát oldatban, ami közelebb áll a fiziológiás körülményekhez. Az NMR képes érzékelni az aminosav oldalláncok atomjainak környezetében bekövetkező változásokat, és ezáltal kimutatni a szubsztrátkötés által indukált mozgásokat. Ez a technika különösen hasznos a gyors, dinamikus folyamatok, például a konformációs átmenetek tanulmányozására.
Fluoreszcencia spektroszkópia
A fluoreszcencia spektroszkópia egy érzékeny módszer, amely a fehérjék belső fluoreszcenciáját (pl. triptofán maradékok) vagy beépített fluoreszcens próbák jeleit használja fel a konformációs változások detektálására. Amikor egy szubsztrát kötődik, az enzim szerkezetében bekövetkező változások módosíthatják a fluoreszcens csoportok környezetét, ami a fluoreszcencia spektrum vagy intenzitás változásában nyilvánul meg.
Krios-elektronmikroszkópia (Cryo-EM)
Az elmúlt években a krios-elektronmikroszkópia (Cryo-EM) forradalmasította a nagy méretű fehérjekomplexek és dinamikus rendszerek szerkezeti vizsgálatát. A Cryo-EM lehetővé teszi a fehérjék különböző konformációs állapotainak vizualizálását, gyakran szubsztráttal vagy anélkül, és így közvetlenül megfigyelhetők az indukált alkalmazkodás során bekövetkező mozgások.
Számítógépes szimulációk és molekuláris dinamika
A számítógépes szimulációk, különösen a molekuláris dinamika (MD) szimulációk, felbecsülhetetlen értékűek az indukált alkalmazkodás mechanizmusának megértésében. Ezek a szimulációk lehetővé teszik a kutatók számára, hogy atomi szinten modellezzék a fehérjék mozgását és kölcsönhatását a szubsztrátokkal időben. Az MD szimulációk révén megfigyelhetővé válnak a konformációs átmenetek, az aktív centrum átrendeződései és a kötődési folyamat dinamikája, megerősítve a kísérleti megfigyeléseket és új betekintést nyújtva a mechanizmusokba.
Ezen technikák kombinációja, a szerkezeti biológia és a biokémia fejlődésével együtt, szilárd alapot biztosít az indukált alkalmazkodási modell érvényességéhez, és folyamatosan bővíti tudásunkat az enzimek dinamikus működéséről.
Az indukált alkalmazkodási modell és a gyógyszertervezés
Az indukált alkalmazkodási modell mélyrehatóan befolyásolta a modern gyógyszertervezés megközelítéseit. A hagyományos gyógyszertervezés gyakran a „zár és kulcs” modellre épült, ahol egy merev célfehérje aktív centrumához tökéletesen illeszkedő molekulát kerestek. Az indukált alkalmazkodás felismerése azonban új dimenziót nyitott meg, hangsúlyozva a fehérjék dinamikus természetét és a ligandum-indukált konformációs változások fontosságát.
A modern gyógyszertervezés ma már figyelembe veszi, hogy a célfehérjék (legyenek azok enzimek, receptorok vagy ioncsatornák) nem statikusak, hanem képesek alkalmazkodni a kötődő molekulákhoz. Ez azt jelenti, hogy egy hatékony gyógyszermolekula nem feltétlenül illeszkedik tökéletesen a célfehérje „üres” aktív centrumához, hanem képes lehet indukálni egy olyan konformációs változást, amely létrehozza az optimális kötőhelyet és/vagy katalitikus aktivitást. Ez a koncepció különösen fontos az allosztérikus gyógyszertervezésben.
Allosztérikus gyógyszerek
Az allosztérikus gyógyszerek nem az aktív centrumba, hanem egy allosztérikus helyre kötődnek, és az indukált alkalmazkodás révén módosítják a célfehérje működését. Ezek a gyógyszerek számos előnnyel járhatnak:
- Nagyobb szelektivitás: Az allosztérikus helyek gyakran kevésbé konzerváltak, mint az aktív centrumok, ami lehetővé teszi a specifikusabb gyógyszerek tervezését, kevesebb mellékhatással.
- Finomabb szabályozás: Az allosztérikus modifikátorok gyakran nem blokkolják teljesen a fehérje működését, hanem modulálják azt, ami finomabb terápiás hatást eredményezhet.
- Telítetlen aktív centrum: Az allosztérikus gyógyszerek nem versenyeznek a természetes szubsztráttal az aktív centrumért, így nagyobb hatékonyságot mutathatnak magas szubsztrátkoncentráció esetén is.
Az indukált alkalmazkodás paradigma-váltást hozott a gyógyszertervezésben, a statikus „zár és kulcs” megközelítésről a dinamikus, rugalmas célpontok felé terelve a figyelmet, megnyitva az utat az allosztérikus gyógyszerek és a precíziós terápia felé.
Virtuális szűrés és de novo tervezés
Az indukált alkalmazkodás figyelembevétele bonyolultabbá teszi a virtuális szűrési (virtual screening) és a de novo gyógyszertervezési (de novo drug design) stratégiákat. Ahelyett, hogy egy merev célfehérjéhez keresnének ligandumokat, a kutatóknak most figyelembe kell venniük a fehérje konformációs rugalmasságát és a ligandum-indukált változásokat. Ez gyakran magában foglalja a „több konformációs állapot” megközelítést (multiple conformation approach), ahol a célfehérje különböző, kísérletileg vagy számítógépesen előállított konformációit használják a szűréshez.
A molekuláris dinamika (MD) szimulációk kulcsfontosságúvá váltak ebben a folyamatban, lehetővé téve a gyógyszerjelöltek kötődésének és a célfehérje konformációs válaszának dinamikus modellezését. Ez segít azonosítani azokat a molekulákat, amelyek nemcsak jól kötődnek, hanem képesek a kívánt konformációs változást is kiváltani.
Receptor-ligandum kölcsönhatások
Az indukált alkalmazkodás elve nem korlátozódik az enzimekre, hanem alapvető fontosságú más receptor-ligandum kölcsönhatásokban is, például a G-protein-kapcsolt receptorok (GPCR-ek) vagy a nukleáris receptorok esetében. Ezek a receptorok a ligandum kötődésekor konformációs változáson mennek keresztül, ami jeltovábbítást vagy génexpressziós változásokat eredményez. A gyógyszerek, amelyek ezeket a receptorokat célozzák, azáltal fejtik ki hatásukat, hogy agonista (aktiváló) vagy antagonista (blokkoló) módon befolyásolják ezeket az indukált konformációs átmeneteket.
Az indukált alkalmazkodási modell tehát nem csupán egy biokémiai elmélet, hanem egy gyakorlati eszköz is, amely forradalmasítja a gyógyszertervezést, lehetővé téve hatékonyabb és specifikusabb terápiák kifejlesztését.
Az indukált alkalmazkodás és a konformációs szelekció: egy continuum

Az indukált alkalmazkodási modell mellett egy másik fontos elmélet is létezik a fehérje-ligandum kölcsönhatások magyarázatára: a konformációs szelekció modell (conformational selection model). Bár elsőre ellentétesnek tűnhetnek, valójában nem kizárólagosak, hanem egy kontinuum két végpontját képviselik, és gyakran együttműködve írják le a valós biológiai rendszerek komplexitását.
Konformációs szelekció modell
A konformációs szelekció modell szerint a fehérjék nem egyetlen, merev konformációban léteznek, hanem dinamikusan egyensúlyban vannak több különböző konformációs állapot között, még ligandum hiányában is. Amikor egy ligandum (pl. szubsztrát) megjelenik, az nem indukál új konformációt, hanem szelektíven kötődik ahhoz a már létező konformációhoz, amelyik a legjobban illeszkedik hozzá. Ezáltal eltolja az egyensúlyt a kötött konformáció irányába.
Képzeljük el, hogy egy fehérje folyamatosan „ingadozik” különböző alakok között. A ligandum egyszerűen „kiválasztja” és stabilizálja azt az alakot, amelyhez a legnagyobb affinitása van. Ez a modell magyarázatot adhat arra, hogy miért képesek egyes enzimek különböző szubsztrátokat kötni, vagy miért lehetnek aktívak és inaktívak is, mielőtt a ligandum megérkezne.
A két modell összehasonlítása és szinergiája
A fő különbség a két modell között a konformációs változás időzítésében és kiváltó okában rejlik:
| Jellemző | Indukált alkalmazkodási modell | Konformációs szelekció modell |
|---|---|---|
| Konformációk létezése | A ligandum (szubsztrát) kötődése indukálja az új, kötött konformációt. | A fehérje már ligandum hiányában is létezik több konformációs állapotban. |
| Kötődés mechanizmusa | A ligandum aktívan formálja a fehérjét a tökéletes illeszkedéshez. | A ligandum szelektíven kötődik egy már létező, optimális konformációhoz. |
| Dinamika hangsúlya | A ligandum-indukált mozgásokra helyezi a hangsúlyt. | A fehérje inherent, ligandum-független dinamikájára helyezi a hangsúlyt. |
A mai tudományos konszenzus szerint a valóság gyakran a két modell közötti kontinuumban helyezkedik el. Sok fehérje valószínűleg mindkét mechanizmust felhasználja valamilyen mértékben. Elképzelhető, hogy egy fehérje már rendelkezik néhány „előformált” konformációval (konformációs szelekció), de a ligandum kötődése még további, finomhangolt változásokat indukálhat (indukált alkalmazkodás) az optimális katalízis vagy funkció elérése érdekében.
A molekuláris szintű kölcsönhatások megértése megköveteli a fehérjék dinamikus természetének felismerését, ahol az indukált alkalmazkodás és a konformációs szelekció gyakran együtt, szinergikusan működnek, hogy a biológiai folyamatok precizitását és hatékonyságát biztosítsák.
Ez a „hibrid” megközelítés lehetővé teszi a kutatók számára, hogy rugalmasabban gondolkodjanak a fehérje-ligandum kölcsönhatásokról, és jobban megértsék a komplex biológiai rendszerek működését. A modern kísérleti technikák és a számítógépes szimulációk egyre jobban képesek feltárni ezeket a finom mechanizmusokat, hozzájárulva a biokémia és a gyógyszerfejlesztés további fejlődéséhez.
Az indukált alkalmazkodás jelentősége a molekuláris biológia szélesebb kontextusában
Az indukált alkalmazkodási modell nem korlátozódik csupán az enzimkatalízisre. Jelentősége sokkal szélesebb körben érvényesül a molekuláris biológia és biokémia különböző területein, alapvető elvként szolgálva a molekuláris kölcsönhatások megértésében.
Receptor-ligandum kölcsönhatások
Ahogy korábban említettük, az indukált alkalmazkodás alapvető a receptor-ligandum kölcsönhatásokban. A sejtfelszíni receptorok, mint például a G-protein-kapcsolt receptorok (GPCR-ek) vagy a tirozinkináz receptorok, a ligandum (pl. hormon, neurotranszmitter) kötődésekor konformációs változáson mennek keresztül. Ez a változás aktiválja a receptor belső tartományát, ami jeltovábbítási kaszkádokat indít el a sejtben, és végső soron sejtes választ eredményez. A ligandum-indukált konformációs változás itt a kulcs a jelátvitelhez.
Fehérje-fehérje kölcsönhatások
A sejtekben a fehérjék ritkán működnek izoláltan. Gyakran komplexeket alkotnak más fehérjékkel, hogy specifikus funkciókat lássanak el. Ezek a fehérje-fehérje kölcsönhatások is gyakran magukban foglalják az indukált alkalmazkodást. Amikor két fehérje kölcsönhatásba lép, mindkét partner konformációs változáson mehet keresztül, hogy optimalizálja a kötődést és a funkcionális interakciót. Ez a mechanizmus biztosítja a fehérjekomplexek összeszerelésének pontosságát és a szabályozás finomhangolását.
Nukleinsav-fehérje kölcsönhatások
A génexpresszió szabályozásában, a DNS replikációjában és a RNS feldolgozásában kulcsfontosságúak a nukleinsav-fehérje kölcsönhatások. A DNS-kötő fehérjék, mint például a transzkripciós faktorok, gyakran indukált alkalmazkodással ismerik fel és kötődnek specifikus DNS-szekvenciákhoz. A fehérje kötődése módosíthatja a DNS szerkezetét (pl. hajlítását), és a DNS is indukálhat konformációs változásokat a fehérjében, hogy optimalizálja a kötődést és a szabályozó funkciót.
Molekuláris gépek működése
Számos biológiai folyamatban, mint például az izomösszehúzódásban, a motorfehérjék működésében vagy a transzportfolyamatokban, komplex molekuláris gépek vesznek részt. Ezek a gépek makromolekuláris komplexek, amelyek összehangolt konformációs változások sorozatán keresztül hajtanak végre mechanikus munkát. Az ATP hidrolízise által szolgáltatott energia gyakran indukált konformációs változásokat idéz elő a molekuláris gépekben, ami lehetővé teszi a mozgást vagy az anyagok szállítását.
Az indukált alkalmazkodás egy univerzális elv, amely a molekuláris kölcsönhatások szívét képezi, lehetővé téve a biológiai rendszerek dinamikus, precíz és adaptív működését a sejten belül és a szervezetekben egyaránt.
Enzimfejlesztés és biokatalízis
Az indukált alkalmazkodás elvének megértése kulcsfontosságú az enzimfejlesztésben és a biokatalízisben. A mérnökileg tervezett enzimek, amelyek új reakciókat katalizálnak vagy megnövelt stabilitással rendelkeznek, gyakran az enzim konformációs rugalmasságának manipulálásával érhetők el. A mesterséges enzimek és a fehérjék tervezése során az indukált alkalmazkodás elveit felhasználva lehet olyan molekulákat létrehozni, amelyek specifikusabbak és hatékonyabbak ipari vagy terápiás alkalmazásokban.
Összességében az indukált alkalmazkodási modell egyike a biokémia legfontosabb elméleteinek, amely alapvetően formálta a molekuláris szintű kölcsönhatásokról alkotott képünket. Dinamikus szemlélete révén jobban megértjük az élő rendszerek rugalmasságát, adaptív képességét és elképesztő precizitását.
Az indukált alkalmazkodás jövőbeli kutatási irányai és kihívásai
Az indukált alkalmazkodási modell alapvető fontosságú a biokémia és a molekuláris biológia számos területén, de még mindig számos nyitott kérdés és kutatási kihívás áll a tudósok előtt. A jövőbeli kutatások valószínűleg a modell finomítására, a mechanizmusok mélyebb megértésére és új alkalmazási területek feltárására fókuszálnak majd.
A konformációs táj feltérképezése
Az egyik legnagyobb kihívás a fehérjék teljes konformációs tájának feltérképezése. A fehérjék nem csupán néhány diszkrét konformáció között váltanak, hanem egy folytonos, sokdimenziós térben mozognak. A modern kísérleti technikák (pl. egyedi molekula fluoreszcencia, nagyfelbontású NMR) és a fejlett számítógépes szimulációk (pl. kiterjesztett MD szimulációk) célja, hogy feltárják ezt a komplex tájat, és azonosítsák azokat az útvonalakat, amelyeken keresztül a fehérjék konformációs változásokat hajtanak végre.
Időfüggő dinamika és a katalízis sebessége
Az indukált alkalmazkodás dinamikus folyamat, amely időben zajlik. A kutatók igyekeznek megérteni, hogy a konformációs változások sebessége hogyan befolyásolja a katalízis sebességét. Melyek a sebességmeghatározó lépések? Mennyire gyorsan képes az enzim alkalmazkodni a szubsztráthoz, és ez hogyan befolyásolja a reakciókinetikát? Az ultragyors spektroszkópiai módszerek és a szabad elektron lézerek (XFEL) ígéretes eszközök lehetnek ezeknek a kérdéseknek a megválaszolására.
Az indukált alkalmazkodás szerepe a betegségekben
Egyre világosabbá válik, hogy az indukált alkalmazkodás zavarai szerepet játszhatnak különböző betegségek kialakulásában. Például, a fehérjék hibás konformációs változásai hozzájárulhatnak a neurodegeneratív betegségekhez (pl. Alzheimer-kór, Parkinson-kór) vagy a rákhoz. A gyógyszerrezisztencia kialakulása is összefüggésbe hozható az enzimek vagy receptorok azon képességével, hogy konformációs változásokkal alkalmazkodnak a gyógyszerekhez, ezáltal hatástalanítva azokat. Ezen mechanizmusok megértése új terápiás stratégiákat nyithat meg.
Racionális enzimtervezés és szintetikus biológia
A jövőben az indukált alkalmazkodás elveit még inkább fel fogják használni a racionális enzimtervezésben. Képesek leszünk-e olyan enzimeket tervezni, amelyek pontosan a kívánt konformációs változást mutatják egy adott szubsztrát jelenlétében, optimalizálva a specifikusságot és a katalitikus hatékonyságot? A szintetikus biológia területén új fehérjéket és molekuláris gépeket hozhatunk létre, amelyek az indukált alkalmazkodás elvét használják fel új funkciók ellátására, például bioszenzorok vagy nanorobotok fejlesztésében.
Az indukált alkalmazkodásról szóló kutatások folyamatosan mélyítik tudásunkat a biológiai rendszerek alapvető működéséről, és ígéretes utakat nyitnak meg a gyógyászat, a biotechnológia és az anyagtudomány területén.
A számítógépes modellezés fejlődése
A számítógépes modellezés, különösen a gépi tanulás és a mesterséges intelligencia (AI) integrálásával, kulcsszerepet fog játszani az indukált alkalmazkodás további megértésében. Az AI-alapú algoritmusok segíthetnek előre jelezni a fehérjék konformációs változásait, optimalizálni a kötődési mechanizmusokat, és felgyorsítani a gyógyszerjelöltek azonosítását. A nagy teljesítményű számítástechnika lehetővé teszi majd a komplexebb rendszerek és hosszabb időskálák szimulálását, közelebb hozva minket a valós biológiai folyamatok atomi szintű megértéséhez.
Az indukált alkalmazkodási modell tehát nem egy lezárt fejezet a tudományban, hanem egy dinamikusan fejlődő terület, amely folyamatosan új felfedezésekkel és innovatív alkalmazásokkal gazdagítja a biokémia és a molekuláris biológia világát.







