
A molekuláris biológia központi dogmája szerint a genetikai információ áramlása egyirányú: a DNS-ből RNS-be, majd RNS-ből fehérjébe. Ezen alapvető folyamat egyik legfontosabb láncszeme a hírvivő RNS, vagy röviden mRNS (messenger RNA). Ez a molekula az élet egyik legcsodálatosabb közvetítője, amely a sejtmagban tárolt genetikai instrukciókat eljuttatja a fehérjeszintézis helyére, a riboszómákhoz. Nélküle a sejtek nem lennének képesek a működésükhöz elengedhetetlen fehérjéket előállítani, így az élet, ahogy ismerjük, értelmezhetetlenné válna.
Az mRNS nem csupán egy egyszerű másolat a DNS-ről; sokkal inkább egy finoman hangolt, dinamikus molekula, amelynek szerkezete és élete szigorúan szabályozott. Képzeljük el úgy, mint egy receptkönyvet, ahol a DNS a teljes szakácskönyv, az mRNS pedig egyetlen receptlap, amelyet a séf (a riboszóma) olvas el, hogy elkészítse az ételt (a fehérjét). Ez a receptlap tele van olyan utasításokkal és jelzésekkel, amelyek befolyásolják, hogy mikor, hol és milyen gyorsan készüljön el az adott étel.
Az elmúlt évtizedekben az mRNS-kutatás hatalmas áttöréseket hozott, különösen az orvostudomány területén. A COVID-19 világjárvány során az mRNS-alapú vakcinák példátlan gyorsasággal történő kifejlesztése és hatékonysága a nagyközönség számára is rávilágított ennek a molekulának a rendkívüli potenciáljára. Ez a cikk célja, hogy mélyrehatóan bemutassa az mRNS szerkezetét, funkcióját és működését, a molekuláris biológia alapjaitól kezdve egészen a modern orvosi alkalmazásokig.
Az RNS típusai és az mRNS helye közöttük
Mielőtt mélyebben belemerülnénk a hírvivő RNS részletes vizsgálatába, érdemes áttekinteni az RNS (ribonukleinsav) molekulák széles családját, hiszen az mRNS csak egy a sok közül, bár kétségtelenül az egyik legfontosabb. Az RNS molekulák alapvetően különböznek a DNS-től (dezoxiribonukleinsavtól) abban, hogy egy ribóz nevű cukormolekulát tartalmaznak dezoxiribóz helyett, és a timin (T) bázis helyett uracil (U) található meg bennük. Bár az RNS gyakran egyszálú, képes komplex háromdimenziós struktúrákat felvenni, amelyek kulcsfontosságúak funkciójuk szempontjából.
Riboszomális RNS (rRNS)
Az rRNS alkotja a riboszómák, a sejtek fehérjeszintetizáló gépezetének strukturális és katalitikus magját. A riboszómák két alegységből állnak, amelyek rRNS-ből és fehérjékből épülnek fel. Az rRNS nemcsak a riboszóma vázát adja, hanem katalizálja a peptidkötések kialakulását is az aminosavak között a fehérjeszintézis során. Ez a katalitikus aktivitás teszi az rRNS-t egyfajta ribozimmé, azaz RNS-enzimmé. Az rRNS a sejt RNS-állományának legnagyobb részét teszi ki.
Transzfer RNS (tRNS)
A tRNS molekulák a fehérjeszintézis (transzláció) során az aminosavakat szállítják a riboszómához az mRNS-en lévő genetikai kódnak megfelelően. Minden tRNS molekula egy specifikus aminosavhoz kapcsolódik, és rendelkezik egy antikodon régióval, amely komplementer az mRNS-en található kodonnal. A tRNS molekulák jellegzetes lóherelevél alakú szerkezettel rendelkeznek, amely tovább hajtogatva egy L-alakú háromdimenziós formát vesz fel, ez esszenciális a funkciójukhoz.
Mikro RNS (miRNS) és egyéb nem kódoló RNS-ek
A nem kódoló RNS-ek (ncRNS-ek) egyre bővülő családjába tartoznak azok az RNS molekulák, amelyek nem kódolnak fehérjéket, de számos fontos szabályozó funkciót látnak el a sejtben. A mikro RNS-ek (miRNS-ek) például rövid, körülbelül 22 nukleotid hosszú RNS-molekulák, amelyek a génexpressziót szabályozzák azáltal, hogy specifikus mRNS-molekulákhoz kapcsolódnak, gátolva azok transzlációját vagy elősegítve lebontásukat. Ezen kívül léteznek hosszú nem kódoló RNS-ek (lncRNS-ek), kis nukleáris RNS-ek (snRNS-ek) és kis nukleoláris RNS-ek (snoRNS-ek), amelyek mind részt vesznek a génexpresszió komplex szabályozásában, az RNS érési folyamatokban és más sejtes funkciókban.
Az mRNS egyedi jellemzői és funkciója
Az mRNS a genetikai információ közvetítője a DNS és a fehérjék között. A DNS-ben tárolt genetikai információt, amely a fehérjék aminosav-szekvenciáját kódolja, az mRNS viszi ki a sejtmagból a citoplazmába, ahol a riboszómák segítségével fehérjékké fordítódik le. Az mRNS egyedi abban, hogy közvetlenül kódolja a fehérjéket, szemben az rRNS és tRNS strukturális és katalitikus szerepével, vagy a nem kódoló RNS-ek szabályozó funkciójával.
Az mRNS molekulák élettartama változatos lehet, a percekben mérhető időtől egészen több óráig, vagy akár napokig is terjedhet, attól függően, hogy milyen fehérjét kódolnak, és milyen gyorsan van szükség a sejtnek az adott fehérje termelésére. Ez a dinamikus szabályozás kulcsfontosságú a sejt adaptációs képességében a változó környezeti feltételekhez.
A hírvivő RNS (mRNS) részletes szerkezete
Az mRNS szerkezete kulcsfontosságú a funkciójához. Bár első pillantásra egyszerűnek tűnhet, mint egy egyszálú nukleinsav, valójában számos finomhangolt elemet tartalmaz, amelyek biztosítják a hatékony és pontos fehérjeszintézist. Ezek az elemek nemcsak a genetikai információ hordozásáért felelősek, hanem a molekula stabilitásáért, szállításáért és a transzláció szabályozásáért is.
Nukleotidok és foszfodiészter kötések: Az RNS alapkövei
Mint minden nukleinsav, az RNS is nukleotidokból épül fel, amelyek ismétlődő egységei polimerizálódnak hosszú lánccá. Minden nukleotid három fő komponensből áll:
- Ribóz cukor: Egy öt szénatomos cukormolekula. A DNS-ben található dezoxiribóztól abban különbözik, hogy a 2′-es szénatomján hidroxilcsoport (-OH) található, míg a dezoxiribóz ezen a pozíción csak hidrogénatomot (-H) tartalmaz. Ez a hidroxilcsoport teszi az RNS-t kémiailag reaktívabbá és kevésbé stabilissá, mint a DNS-t.
- Foszfát csoport: Negatív töltésű csoport, amely a cukor 5′-ös szénatomjához kapcsolódik. A foszfát csoportok képezik az RNS gerincét, összekapcsolva az egyes nukleotidokat.
- Nitrogéntartalmú bázis: Négyféle bázis létezik az RNS-ben:
- Adenin (A)
- Guanin (G)
- Citozin (C)
- Uracil (U)
Fontos különbség a DNS-hez képest, hogy az RNS-ben a timin (T) helyett uracil (U) található. Az adenin és a guanin purin bázisok (kétgyűrűs szerkezetűek), míg a citozin és az uracil pirimidin bázisok (egygyűrűs szerkezetűek).
Az egyes nukleotidok egymáshoz foszfodiészter kötésekkel kapcsolódnak. Ez a kötés a ribóz 3′-ös szénatomján lévő hidroxilcsoport és a következő nukleotid 5′-ös szénatomjához kapcsolódó foszfátcsoport között alakul ki. Ennek eredményeként egy hosszú, irányított polimer lánc jön létre, amelynek egyik vége 5′-foszfát csoporttal, a másik vége 3′-hidroxil csoporttal végződik. Ez az 5′ → 3′ polaritás alapvető fontosságú a genetikai információ leolvasása szempontjából.
Egyszálú természet: Miért fontos ez?
Bár a DNS jellemzően kétszálú hélixet alkot, az mRNS általában egyszálú. Ez az egyszálú természet lehetővé teszi számára, hogy flexibilis legyen, és komplex háromdimenziós struktúrákat vegyen fel azáltal, hogy a lánc különböző részei egymással komplementer bázispárosodásokat alakítanak ki (pl. hajtűhurkok, hurkok). Ezek a másodlagos és harmadlagos szerkezetek kritikusak az mRNS stabilitása, a fehérjékhez való kötődése és a transzláció szabályozása szempontjából. Az egyszálú jelleg egyben azt is jelenti, hogy az mRNS sokkal hozzáférhetőbb a riboszómák és más RNS-kötő fehérjék számára, mint egy kétszálú DNS molekula.
Az mRNS specifikus régiói
Az mRNS molekula nem csupán a fehérjét kódoló szekvenciából áll. Számos nem kódoló régiót is tartalmaz, amelyek létfontosságúak a molekula életciklusában, a génexpresszió szabályozásában és a fehérjeszintézis hatékonyságában. Ezek a régiók az 5′-végen és a 3′-végen helyezkednek el, és kulcsszerepet játszanak a molekula stabilitásában, szállításában és a transzláció szabályozásában.
5′ sapka (5′ cap)
Az eukarióta mRNS molekulák 5′-vége egy speciális módosítást hordoz, az úgynevezett 5′ sapkát (5′ cap). Ez egy 7-metilguanozin molekula, amely egy szokatlan 5′-5′ trifoszfát kötéssel kapcsolódik az mRNS első nukleotidjához. Az 5′ sapka számos fontos funkciót lát el:
- Védelmet nyújt: Megvédi az mRNS-t a 5′ irányú exonukleázok általi lebontástól, növelve a molekula stabilitását.
- Riboszóma felismerés: Elengedhetetlen a riboszóma kis alegységének felismeréséhez és az mRNS-hez való kötődéséhez a transzláció iniciációja során. A sapka egyfajta „felismerő jel” a fehérjeszintetizáló gépezet számára.
- Export a sejtmagból: Segít az mRNS sejtmagból a citoplazmába történő exportálásában.
A sapka hozzáadása a transzkripció korai szakaszában történik, még a sejtmagban, az RNS polimeráz II által.
5′ nem transzlálódó régió (5′ UTR)
Az 5′ nem transzlálódó régió (5′ UTR) az 5′ sapka és a fehérjét kódoló szekvencia (CDS) között helyezkedik el. Ez a régió nem fordítódik le fehérjévé, de rendkívül fontos a transzláció szabályozásában. Tartalmazhat olyan szekvenciákat, amelyek:
- Szabályozó fehérjék kötőhelyei: Különböző RNS-kötő fehérjék (RBP-k) kapcsolódhatnak ide, befolyásolva a transzláció hatékonyságát, vagy akár gátolva azt.
- Másodlagos szerkezetek: Komplex hajtűhurkokat vagy egyéb szerkezeteket képezhet, amelyek befolyásolják a riboszóma hozzáférését a start kodonhoz.
Kódoló régió (CDS)
A kódoló régió (Coding Sequence, CDS) az mRNS azon része, amely a fehérje aminosav-szekvenciáját kódolja. Ez a régió a start kodonnal (általában AUG) kezdődik és egy stop kodonnal (UAA, UAG, vagy UGA) végződik. A start kodon jelzi a riboszómának, hogy hol kezdje el a fehérjeszintézist, míg a stop kodon jelzi a befejezést. A CDS-ben található kodonok sorrendje határozza meg a szintetizálandó fehérje aminosav-szekvenciáját.
3′ nem transzlálódó régió (3′ UTR)
A 3′ nem transzlálódó régió (3′ UTR) a stop kodon után, de a poli(A) farok előtt helyezkedik el. Ez a régió is kulcsszerepet játszik az mRNS sorsának szabályozásában, beleértve a stabilitást, a lokalizációt és a transzláció hatékonyságát. Fontos funkciói:
- RNS-kötő fehérjék kötőhelyei: Számos RBP köthető ide, amelyek befolyásolják az mRNS élettartamát és transzlációját.
- MikroRNS-ek célpontja: A miRNS-ek gyakran a 3′ UTR-hez kötődnek, gátolva a transzlációt vagy elősegítve az mRNS lebontását.
- Lokalizációs szekvenciák: Bizonyos mRNS-ek specifikus sejtterületekre való szállítását irányító szekvenciákat tartalmazhat.
A 3′ UTR komplexitása és szabályozó potenciálja miatt a génexpresszió poszt-transzkripciós szabályozásának egyik legfontosabb eleme.
Poli(A) farok
Az eukarióta mRNS molekulák 3′-végéhez egy hosszú, több száz adenin nukleotidból álló szakasz, a poli(A) farok kapcsolódik. Ezt a farokrészt a poli(A) polimeráz enzim adja hozzá a transzkripció befejezése után, függetlenül a DNS templátjától. A poli(A) faroknak több fontos funkciója van:
- Stabilitás növelése: Védi az mRNS-t a 3′ irányú exonukleázok általi lebontástól. Minél hosszabb a poli(A) farok, annál stabilabb az mRNS általában.
- Transzláció elősegítése: Segít a riboszómának az mRNS-hez való kötődésében és a transzláció iniciációjában. Kölcsönhatásba léphet az 5′ sapkával, kör alakú mRNS struktúrát képezve, ami hatékonyabbá teszi a transzlációt.
- Export a sejtmagból: Részt vesz az mRNS sejtmagból való kijutásában.
A poli(A) farok hossza dinamikusan változhat, és a farok rövidülése gyakran jelzi az mRNS lebontásának kezdetét.
Az mRNS szerkezete nem csupán a genetikai üzenet hordozója, hanem egy kifinomult szabályozó platform, amely biztosítja a precíz és hatékony fehérjeszintézist.
A genetikai kód és az mRNS kapcsolata
A genetikai kód az a szabályrendszer, amely meghatározza, hogyan fordítódik le az mRNS nukleotid-szekvenciája egy fehérje aminosav-szekvenciájává. Ez a kód egyetemes, ami azt jelenti, hogy szinte minden élőlényben ugyanazokat a szabályokat követi, a baktériumoktól az emberig. Az mRNS a genetikai kód hordozója, és a rajta lévő információt a riboszómák és a tRNS molekulák olvaszák le.
A kodonok rendszere
A genetikai kód alapvető egysége a kodon, amely három egymást követő nukleotidból álló szekvencia az mRNS-en. Minden kodon egy specifikus aminosavat kódol, vagy jelzi a fehérjeszintézis kezdetét vagy végét. Mivel négyféle nukleotid létezik (A, U, G, C), és ezek hármas csoportokban kombinálódnak, elméletileg 43 = 64 különböző kodon lehetséges.
Az aminosavak száma azonban mindössze 20. Ez azt jelenti, hogy több kodon is kódolhatja ugyanazt az aminosavat. Ezt a jelenséget nevezzük a genetikai kód degeneráltságának vagy redundanciájának. Például a leucin aminosavat hat különböző kodon is kódolhatja (UUA, UUG, CUU, CUC, CUA, CUG). Ez a degeneráltság fontos védelmi mechanizmus a mutációk ellen, mivel egyetlen nukleotid megváltozása nem feltétlenül vezet az aminosav megváltozásához, és így a fehérje funkciójának elvesztéséhez.
Start és stop kodonok
Az mRNS-en található kodonok között különleges szerepük van a start és stop kodonoknak:
- Start kodon: Az AUG kodon a leggyakoribb start kodon, amely a metionin (Met) aminosavat kódolja. Ezen kívül jelzi a riboszómának, hogy hol kezdje el a transzlációt. Az eukariótákban a metionin általában egy speciális iniciátor tRNS-sel kerül beépítésre.
- Stop kodonok: Három stop kodon létezik: UAA, UAG és UGA. Ezek a kodonok nem kódolnak egyetlen aminosavat sem. Ehelyett azt jelzik a riboszómának, hogy fejezze be a fehérjeszintézist. Amikor a riboszóma egy stop kodonhoz ér, felszabadító faktorok kötődnek hozzá, és a szintetizált polipeptidlánc leválik a riboszómáról.
A genetikai kód egyetemessége
A genetikai kód egyetemessége az egyik legmegdöbbentőbb felfedezés a molekuláris biológiában. Ez azt jelenti, hogy az AUG kodon szinte minden élőlényben metionint kódol, és a stop kodonok is univerzálisan a szintézis leállítását jelzik. Ez az egyetemesség erőteljes bizonyítékot szolgáltat a földi élet közös eredetére. Bár vannak apró eltérések (pl. mitokondriális genetikai kód), az alapvető szabályrendszer mindenhol ugyanaz.
Az mRNS a genetikai kód közvetítőjeként tehát egy rendkívül precíz és komplex információátviteli folyamat kulcsfontosságú szereplője. A kodonok sorrendje az mRNS-en határozza meg a fehérje primér szerkezetét, amely végső soron a fehérje háromdimenziós alakját és funkcióját is meghatározza.
Az mRNS funkciója: a fehérjeszintézis vezérlése

Az mRNS elsődleges funkciója a fehérjeszintézis vezérlése. Ez a folyamat két fő lépésből áll: a transzkripcióból, ahol a DNS-ről RNS másolat készül, és a transzlációból, ahol az mRNS információja alapján fehérje szintetizálódik. Ez a két lépés alkotja a molekuláris biológia központi dogmáját, amely az élet alapvető információáramlási útvonalát írja le.
Transzkripció: a DNS-ről az mRNS-re
A transzkripció az a folyamat, amelynek során a DNS genetikai információja átíródik egy RNS molekulába. Eukarióta sejtekben ez a sejtmagban történik. A folyamat főszereplője az RNS polimeráz enzim.
Az RNS polimeráz szerepe
Az RNS polimeráz egy enzim, amely katalizálja az RNS szál szintézisét a DNS templát alapján. Nem igényel primer-t, ellentétben a DNS polimerázzal. Az RNS polimeráz a DNS kettős spirálját feltekeri, az egyik DNS szálat templátként használja, és komplementer RNS nukleotidokat illeszt be a növekvő RNS láncba. A DNS-ben lévő adeninhez uracil, a guaninhez citozin, a citozinhez guanin és a timinhez adenin kapcsolódik.
Eukariótákban három fő RNS polimeráz létezik:
- RNS polimeráz I: rRNS-t szintetizál.
- RNS polimeráz II: mRNS-t, snRNS-t és miRNS-t szintetizál. Ez a legfontosabb az mRNS képzés szempontjából.
- RNS polimeráz III: tRNS-t és egyéb kis RNS-eket szintetizál.
Promóter és terminátor régiók
A transzkripció nem véletlenszerűen kezdődik a DNS-en. Az RNS polimeráz specifikus DNS szekvenciákhoz, az úgynevezett promóter régiókhoz kötődik. A promóterek jelzik a gén kezdetét és irányítják az RNS polimeráz kötődését. Különböző transzkripciós faktorok segítik az RNS polimerázt a promóter felismerésében és a transzkripció megkezdésében.
A transzkripció a terminátor régiók elérésével fejeződik be. Ezek a szekvenciák jelzik az RNS polimeráznak, hogy váljon le a DNS-ről és szabadítsa fel az újonnan szintetizált RNS molekulát.
Eukarióta és prokarióta transzkripció különbségei
A transzkripció folyamata lényegében hasonló a prokariótákban és az eukariótákban, de vannak jelentős különbségek:
| Jellemző | Prokarióták | Eukarióták |
|---|---|---|
| Helyszín | Citoplazma | Sejtmag |
| RNS polimerázok | Egy típus | Három típus (Pol I, II, III) |
| Pre-mRNS érés | Nincs (policisztás mRNS) | Komplex érés (splicing, capping, poliadeniláció) |
| Transzkripció és transzláció | Egyszerre történhet (kuplungolt) | Térben és időben elkülönül |
| Génstruktúra | Folyamatos kódoló régiók | Intronok és exonok |
Elő-mRNS (pre-mRNA) és az érési folyamatok
Eukarióta sejtekben az RNS polimeráz II által szintetizált elsődleges transzkriptumot elő-mRNS-nek (pre-mRNA) nevezzük. Ez az elő-mRNS még nem funkcionális mRNS, és számos érésen (processing) kell keresztülmennie a sejtmagban, mielőtt a citoplazmába kerülhetne transzlációra. Ezek a módosítások:
- 5′ sapka hozzáadása (Capping): Ahogy már említettük, egy 7-metilguanozin molekula kapcsolódik az mRNS 5′-végéhez egy szokatlan 5′-5′ trifoszfát kötéssel. Ez a transzkripció korai szakaszában történik.
- Splicing (Intronok eltávolítása): Az eukarióta gének gyakran tartalmaznak intronokat (nem kódoló szekvenciák) és exonokat (kódoló szekvenciák). A splicing során az intronokat kivágják, és az exonokat pontosan összekapcsolják. Ezt a folyamatot a spliceoszóma végzi, amely snRNS-ekből és fehérjékből áll. A splicing lehetővé teszi az alternatív splicingot is, ahol egyetlen génből különböző mRNS izoformák és ezáltal különböző fehérjék keletkezhetnek.
- Poliadeniláció (Poli(A) farok hozzáadása): A transzkripció befejezése után egy poli(A) farok (több száz adenin nukleotidból álló szakasz) kerül hozzáadásra az mRNS 3′-végéhez. Ezt a poli(A) polimeráz enzim végzi.
Ezek az érési lépések biztosítják, hogy csak a funkcionális, érett mRNS molekulák jussanak ki a sejtmagból, és hatékonyan transzlalódjanak fehérjékké.
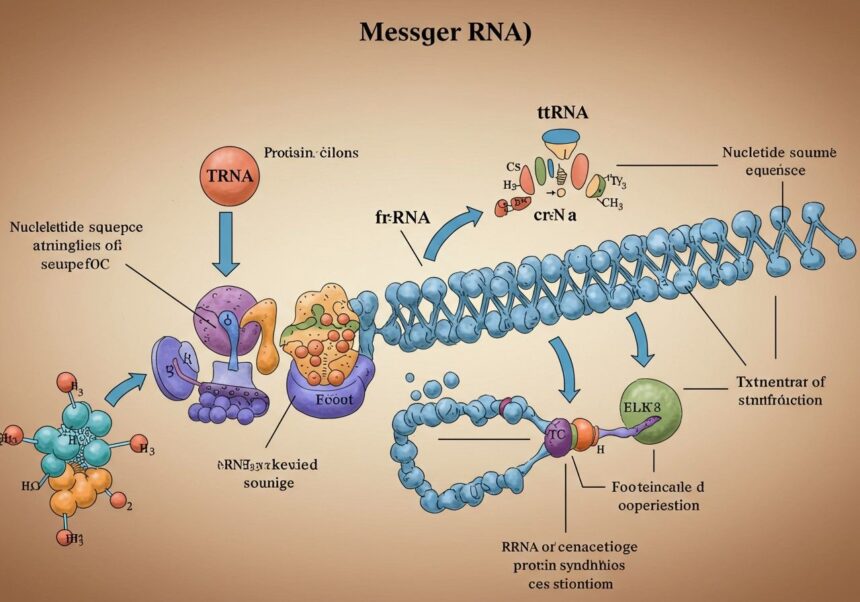
Transzláció: az mRNS-ről a fehérjére
A transzláció az a folyamat, amelynek során az mRNS-en lévő genetikai információ átalakul egy aminosav-szekvenciává, azaz egy fehérjévé. Ez a citoplazmában történik, a riboszómák, a tRNS molekulák és számos fehérje faktor részvételével.
A riboszómák szerkezete és funkciója
A riboszómák a sejtek fehérjeszintetizáló gyárai. Két alegységből állnak: egy nagy és egy kis alegységből, amelyek rRNS-ből és fehérjékből épülnek fel. A riboszóma feladata, hogy az mRNS-hez kötődjön, leolvassa a kodonokat, és megfelelő tRNS-ek segítségével összekapcsolja az aminosavakat peptidkötésekkel, létrehozva a növekvő polipeptidláncot.
A riboszómán belül három tRNS kötőhely található:
- A-hely (Aminoacil-tRNS hely): Ide kötődik az új, aminosavval feltöltött tRNS.
- P-hely (Peptidil-tRNS hely): Ide kötődik a tRNS, amely a növekvő polipeptidláncot hordozza.
- E-hely (Exit hely): Innen távozik a már aminosav nélküli tRNS.
A tRNS szerepe: antikodon és aminosav kötés
A tRNS molekulák a „fordítók” szerepét töltik be a transzláció során. Minden tRNS egy specifikus aminosavat képes megkötni a 3′-végén. Ezen kívül rendelkezik egy antikodon régióval, amely három nukleotidból áll, és komplementer bázispárosodással kapcsolódik az mRNS-en lévő kodonhoz. Ez a komplementaritás biztosítja, hogy a megfelelő aminosav kerüljön beépítésre a növekvő fehérjeláncba.
A transzláció fázisai: iniciáció, elongáció, termináció
A transzláció három fő fázisra osztható:
- Iniciáció (Indítás): Ez a fázis a riboszóma, az mRNS és az első aminosavval (metionin) töltött iniciátor tRNS összeállásával kezdődik. Eukariótákban az iniciátor tRNS először a riboszóma kis alegységéhez kötődik, majd ez a komplex az mRNS 5′ sapkájához kapcsolódik. Ezután elkezdi szkennelni az mRNS-t az 5′ UTR-en keresztül, amíg el nem éri az AUG start kodont. Ekkor a nagy riboszóma alegység is csatlakozik, és az iniciátor tRNS a P-helyre kerül.
- Elongáció (Láncnövelés): Az elongáció során a riboszóma lépésről lépésre halad az mRNS-en (5′ → 3′ irányban), és aminosavakat ad hozzá a növekvő polipeptidlánchoz.
- Egy új, aminosavval töltött tRNS érkezik az A-helyre, amelynek antikodonja komplementer az mRNS aktuális kodonjával.
- A riboszóma katalizálja a peptidkötés kialakulását a P-helyen lévő tRNS által hordozott polipeptidlánc és az A-helyen lévő új aminosav között.
- A riboszóma ezután elmozdul egy kodonnyit (transzlokáció). Ezzel a P-helyen lévő tRNS (most már a teljes polipeptidlánccal) az E-helyre kerül, az A-helyen lévő tRNS (most már a polipeptidlánc végével) a P-helyre kerül, és az A-hely szabaddá válik a következő aminosavval töltött tRNS számára. Az E-helyről az aminosav nélküli tRNS távozik.
Ez a ciklus ismétlődik, amíg a riboszóma el nem ér egy stop kodont.
- Termináció (Befejezés): Amikor a riboszóma egy stop kodonhoz (UAA, UAG, UGA) ér az mRNS-en, nem kötődik hozzá tRNS. Ehelyett felszabadító faktorok (release factors) kötődnek az A-helyre. Ezek a faktorok katalizálják a polipeptidlánc hidrolízisét a tRNS-ről, és a frissen szintetizált fehérje leválik a riboszómáról. Ezt követően a riboszóma alegységei szétválnak, és készen állnak egy új transzlációs ciklusra.
Poliszómák
Gyakran előfordul, hogy több riboszóma is transzlalja ugyanazt az mRNS molekulát egyidejűleg. Ezeket a struktúrákat poliszómáknak vagy poliriboszómáknak nevezzük. A poliszómák lehetővé teszik a sejt számára, hogy gyorsan és hatékonyan termeljen nagy mennyiségű fehérjét egyetlen mRNS molekula felhasználásával, maximalizálva a transzlációs kapacitást.
Az mRNS a genetikai információ hídja, amely a DNS-ben rejlő potenciált aktív, működő fehérjékké alakítja, az élet minden alapvető folyamatához nélkülözhetetlenül.
Az mRNS stabilitása és lebontása
Az mRNS molekulák nem örökkévalóak. Élettartamuk szigorúan szabályozott, ami alapvető fontosságú a génexpresszió finomhangolásában. A sejtnek pontosan tudnia kell, mikor van szüksége egy adott fehérjére, és mikor kell leállítani annak termelését. Az mRNS stabilitása, azaz az a sebesség, amellyel lebomlik, döntő mértékben befolyásolja a fehérjeszintézis mértékét. Egy hosszú élettartamú mRNS sok fehérjét termelhet, míg egy rövid élettartamú mRNS csak keveset.
Az mRNS élettartamának szabályozása
Az mRNS élettartamának szabályozása egy komplex folyamat, amely számos tényezőtől függ, beleértve:
- A gén típusa: Például a stresszre reagáló fehérjéket kódoló mRNS-ek gyakran rövid élettartamúak, hogy a sejt gyorsan alkalmazkodhasson.
- A sejt típusa és fejlődési állapota: Különböző sejtekben és fejlődési fázisokban eltérő lehet az mRNS stabilitása.
- Környezeti jelek: Hormonok, növekedési faktorok vagy stresszhatások mind befolyásolhatják az mRNS stabilitását.
- Az mRNS specifikus szekvenciái és szerkezetei: Különösen az UTR régiókban lévő elemek.
Enzimek (RNázok) szerepe
Az mRNS lebontását speciális enzimek, az úgynevezett RNázok (ribonukleázok) végzik. Ezek az enzimek képesek az RNS lánc foszfodiészter kötéseit hasítani, darabokra bontva a molekulát. Két fő típusuk van:
- Exonukleázok: Az RNS szál végeiről, vagy az 5′-ről vagy a 3′-ről haladva távolítják el a nukleotidokat.
- Endonukleázok: Az RNS szál belső részén hasítják el a foszfodiészter kötéseket.
A lebontás gyakran a poli(A) farok rövidülésével (deadeniláció) kezdődik, amelyet a 3′-ről történő lebontás vagy az 5′ sapka eltávolítása (decapping), majd az 5′-ről történő lebontás követ.
A stabilitást befolyásoló tényezők
Az mRNS stabilitását számos tényező befolyásolja, amelyek kölcsönhatásban állnak egymással:
- Poli(A) farok hossza: A poli(A) farok hossza közvetlenül korrelál az mRNS stabilitásával. A poli(A) farok rövidülése (deadeniláció) gyakran az első lépés az mRNS lebontása felé. A poli(A) farok hosszát poli(A) polimerázok (hosszabbítás) és deadenilázok (rövidítés) szabályozzák.
- 5′ sapka: Az 5′ sapka védelmet nyújt az 5′ exonukleázok ellen. A sapka eltávolítása (decapping) gyorsan vezet az mRNS lebontásához.
- UTR-ek (nem transzlálódó régiók): Az 5′ UTR és a 3′ UTR gyakran tartalmaznak cisz-hatású elemeket (specifikus nukleotid szekvenciákat), amelyekhez transz-hatású faktorok (RNS-kötő fehérjék, RBP-k) kapcsolódhatnak. Ezek az RBP-k befolyásolhatják az mRNS stabilitását, akár növelve, akár csökkentve azt. Például, az AT-gazdag régiók (ARE-k) a 3′ UTR-ben gyakran instabil mRNS-eket jelölnek.
- MikroRNS-ek (miRNS-ek): Ahogy már említettük, a miRNS-ek a 3′ UTR-hez kötődhetnek, és az RISC (RNA-induced silencing complex) komplex segítségével gátolhatják a transzlációt, vagy elősegíthetik az mRNS lebontását.
A minőség-ellenőrzési mechanizmusok (NMD – Nonsense-mediated decay)
A sejtek rendkívül kifinomult minőség-ellenőrzési mechanizmusokkal rendelkeznek, amelyek biztosítják, hogy csak hibátlan mRNS molekulák termeljenek fehérjéket, és a hibás mRNS-ek gyorsan lebomoljanak. Az egyik legfontosabb ilyen mechanizmus a nonsense-mediated decay (NMD).
Az NMD felismeri és lebontja azokat az mRNS molekulákat, amelyek idő előtti stop kodont (premature termination codon, PTC) tartalmaznak. Ezek a PTC-k gyakran mutációk vagy hibás splicing eredményeként jönnek létre, és csonka, diszfunkcionális fehérjék termeléséhez vezetnének. Az NMD mechanizmus biztosítja, hogy az ilyen hibás mRNS-ek ne halmozódjanak fel, ezzel védve a sejtet a káros fehérjék termelésétől és fenntartva a genetikai integritást.
Az mRNS szerepe a génexpresszió szabályozásában
A génexpresszió szabályozása egy rendkívül komplex folyamat, amely biztosítja, hogy a megfelelő gének a megfelelő időben és a megfelelő mennyiségben fejeződjenek ki. Az mRNS nem csupán passzív hordozója a genetikai információnak, hanem aktív résztvevője is ennek a szabályozásnak, különösen a poszt-transzkripciós szinten.
Transzkripciós és poszt-transzkripciós szabályozás
A génexpresszió szabályozása számos szinten történhet:
- Transzkripciós szint: Ez a szabályozás a DNS-ről RNS-re történő átírás folyamatát befolyásolja. Transzkripciós faktorok, cisz-hatású elemek (pl. promóterek, enhanszerek) és epigenetikai módosítások (pl. DNS metiláció, hiszton módosítások) mind hozzájárulnak ahhoz, hogy egy gén aktiválódjon vagy elhallgattassék.
- Poszt-transzkripciós szint: Ez a szabályozás az RNS molekula érését, stabilitását, szállítását és transzlációját érinti. Az mRNS-nek kulcsszerepe van ezen a szinten, mivel számos mechanizmus befolyásolja a sorsát a transzkripció után.
RNS-kötő fehérjék (RBP-k)
Az RNS-kötő fehérjék (RBP-k) egy sokszínű fehérjecsalád, amelyek specifikus RNS szekvenciákhoz vagy szerkezetekhez kötődnek. Ezek a fehérjék rendkívül fontosak az mRNS életciklusának minden szakaszában, a transzkripciótól a lebontásig. Az RBP-k befolyásolhatják:
- Splicing: Szabályozzák az intronok kivágását és az exonok összekapcsolását.
- mRNS stabilitás: Egyes RBP-k védik az mRNS-t a lebontástól, míg mások elősegítik azt.
- Transzláció hatékonysága: Befolyásolhatják a riboszóma kötődését az mRNS-hez és a fehérjeszintézis sebességét.
- mRNS lokalizáció: Irányíthatják az mRNS-t specifikus sejtterületekre, biztosítva, hogy a fehérjék ott szintetizálódjanak, ahol szükség van rájuk.
- Poli(A) farok szabályozása: Befolyásolják a poli(A) farok hosszát és dinamikáját.
Az RBP-k és az mRNS közötti komplex kölcsönhatások hálózata finomhangolja a génexpressziót, lehetővé téve a sejt számára, hogy gyorsan és pontosan reagáljon a belső és külső ingerekre.
Nem kódoló RNS-ek (ncRNS-ek) és az mRNS kölcsönhatásai
A nem kódoló RNS-ek, különösen a mikroRNS-ek (miRNS-ek), jelentős szabályozó szerepet töltenek be az mRNS-sel való kölcsönhatásuk révén. A miRNS-ek rövid, egyszálú RNS molekulák, amelyek komplementer bázispárosodással kötődnek az mRNS 3′ UTR régiójához.
Ez a kötődés általában a következő következményekkel jár:
- Transzláció gátlása: A miRNS-RISC komplex fizikai akadályt képezhet a riboszóma számára, vagy befolyásolhatja az iniciációs faktorokat, ezáltal csökkentve a fehérjeszintézis mértékét.
- mRNS lebontás elősegítése: A miRNS-RISC komplex toborozhat olyan enzimeket (deadenilázokat, decapping enzimeket, exonukleázokat), amelyek felgyorsítják az mRNS lebontását.
A miRNS-ek egyetlen mRNS-hez, vagy akár több különböző mRNS-hez is kötődhetnek, és egyetlen mRNS-t több miRNS is szabályozhat. Ez a hálózat rendkívül komplex és precíz szabályozást tesz lehetővé, ami alapvető a sejtfejlődésben, differenciációban és a betegségek kialakulásában.
Ezen kívül, más ncRNS-ek, mint például a hosszú nem kódoló RNS-ek (lncRNS-ek), szintén kölcsönhatásba léphetnek az mRNS-sel, befolyásolva annak stabilitását, szállítását vagy transzlációját. Az lncRNS-ek működési mechanizmusai sokfélék lehetnek, például RBP-ket toborozhatnak az mRNS-hez, vagy akár fizikai scaffoldként (állványként) is szolgálhatnak komplexek számára.
Az mRNS nem csupán egy tervrajz; egy dinamikus és szabályozott molekula, amely aktívan részt vesz a génexpresszió finomhangolásában, meghatározva a sejt sorsát és működését.
Az mRNS technológiai és orvosi alkalmazásai
Az mRNS molekuláris biológiai alapjainak megértése hatalmas lehetőségeket nyitott meg a biotechnológia és az orvostudomány számára. Az mRNS-alapú technológiák az elmúlt évtizedben forradalmi áttöréseket hoztak, különösen a vakcinológia és a terápiás megközelítések terén. A COVID-19 világjárvány során az mRNS vakcinák sikere katapultálta ezt a technológiát a köztudatba, de alkalmazási területei messze túlmutatnak a fertőző betegségeken.
mRNS vakcinák
Az mRNS vakcinák forradalmasították a fertőző betegségek elleni védekezést. Működési elvük rendkívül elegáns és hatékony:
Működési elv (antigén termelés)
A hagyományos vakcinák inaktivált vírusokat, legyengített kórokozókat vagy tisztított fehérjéket tartalmaznak, amelyek az immunrendszert antitestek termelésére serkentik. Az mRNS vakcinák ezzel szemben nem tartalmaznak magát a kórokozót. Ehelyett egy mRNS molekulát juttatnak be a szervezetbe, amely egy specifikus vírusfehérjét (antigént) kódol, például a SARS-CoV-2 vírus tüskefehérjéjét. Az oltás beadása után a sejtek felveszik az mRNS-t, és a sejt saját riboszómái felhasználásával lefordítják azt a vírusfehérjévé. Ez a fehérje megjelenik a sejtek felszínén, vagy felszabadul a sejtből, és az immunrendszer felismeri, mintha egy valós fertőzés lenne. Ennek hatására az immunrendszer antitesteket és T-sejteket termel, amelyek felkészülnek a jövőbeli fertőzés elleni védekezésre.
Előnyök
- Gyors fejlesztés és gyártás: Mivel az mRNS vakcinák szintetikus úton, sejtkultúrák helyett laboratóriumban állíthatók elő, fejlesztésük és gyártásuk sokkal gyorsabb, mint a hagyományos vakcináké. Ez kritikus fontosságú volt a gyorsan terjedő világjárvány idején.
- Biztonságosság: Az mRNS nem jut be a sejtmagba, és nem integrálódik a gazdaszervezet genomjába, így nincs kockázata a genetikai módosulásnak. Az mRNS molekula rövid élettartamú, és a fehérje termelése után lebomlik.
- Erős immunválasz: Az mRNS vakcinák általában erős humorális (antitest) és celluláris (T-sejt) immunválaszt váltanak ki.
- Skálázhatóság: A gyártási folyamat viszonylag könnyen skálázható nagy mennyiségű vakcina előállítására.
- Nem tartalmaz adjuvánst: Sok hagyományos vakcina adjuvánsokat (immunválaszt erősítő anyagokat) tartalmaz, az mRNS vakcinák általában nem igényelnek ilyet, mivel az mRNS önmagában is képes immunválaszt kiváltani.
Kihívások
- Stabilitás: Az mRNS molekula rendkívül instabil és érzékeny az RNázok általi lebontásra. Ezért speciális szállítási rendszerekre és tárolási körülményekre (pl. ultra-hideg hőmérséklet) van szükség.
- Szállítás: Az mRNS-nek be kell jutnia a sejtekbe, de a szabad mRNS könnyen lebomlik és nem jut át a sejtmembránon. Ezért lipid nanorészecskékbe (LNP-k) csomagolják, amelyek védelmet nyújtanak és elősegítik a sejtekbe való felvételt.
COVID-19 vakcinák sikere
A Pfizer-BioNTech (Comirnaty) és a Moderna (Spikevax) mRNS vakcinák példátlan sikert arattak a COVID-19 világjárvány elleni küzdelemben. Hatalmas hatékonyságot mutattak a súlyos betegség, a kórházi kezelés és a halálesetek megelőzésében. Ez a siker nemcsak a pandémia megfékezésében játszott kulcsszerepet, hanem bebizonyította az mRNS technológia életképességét és hatalmas potenciálját a jövőbeli orvosi alkalmazások számára.
Rákterápia
Az mRNS technológia ígéretes utakat nyit meg a rákterápia területén is. Az mRNS-t fel lehet használni:
- Rákellenes vakcinák: Az mRNS kódolhatja a rákos sejtekre jellemző antigéneket (tumor-specifikus antigének). Ezek az antigének bemutatása az immunrendszernek rákellenes immunválaszt válthat ki, amely megtámadja és elpusztítja a tumorsejteket. Ez a megközelítés személyre szabott rákvakcinák fejlesztését is lehetővé teheti, ahol a beteg saját tumorából származó mutációkat kódoló mRNS-t használnak.
- Immunitás stimulálása: Az mRNS kódolhat immunszabályozó molekulákat, például citokineket, amelyek helyileg stimulálják az immunrendszert a tumor mikro környezetében, fokozva az immunterápia hatékonyságát.
Génterápia
A génterápia célja a genetikai betegségek gyógyítása a hibás gének korrigálásával vagy pótlásával. Az mRNS alapú génterápia számos előnnyel jár:
- Hiányzó fehérjék pótlása: Az mRNS bejuttatható a sejtekbe, hogy ideiglenesen termeljen egy hiányzó vagy hibás fehérjét. Ez különösen hasznos lehet olyan betegségek esetén, mint a cisztás fibrózis, ahol egy specifikus fehérje (CFTR) hiánya okozza a problémát.
- Genomszerkesztés elősegítése: Az mRNS használható a génszerkesztő eszközök (pl. CRISPR/Cas9) szállítására a sejtekbe, ahol azok módosíthatják a DNS-t. Mivel az mRNS átmeneti, csökkenti a nem kívánt off-target hatások kockázatát, ellentétben a DNS alapú génszerkesztő vektorokkal.
Regeneratív medicina
A regeneratív medicina célja a sérült szövetek és szervek helyreállítása vagy pótlása. Az mRNS ebben is szerepet játszhat:
- Sejtek átprogramozása: Az mRNS használható a sejtek átprogramozására, például fibroblasztokból indukált pluripotens őssejtek (iPSC-k) létrehozására, amelyek aztán differenciálhatók különböző sejttípusokká. Ez a módszer biztonságosabb, mint a vírusvektorok alkalmazása, mivel az mRNS nem integrálódik a genomba.
- Növekedési faktorok termelése: Az mRNS kódolhat olyan növekedési faktorokat, amelyek serkentik a szövetek regenerációját és sebgyógyulását.
Diagnosztika
Az mRNS-t diagnosztikai célokra is fel lehet használni, például:
- Biomarkerek azonosítása: Bizonyos betegségek, például a rák vagy fertőző betegségek esetén specifikus mRNS molekulák expressziós mintázata változhat. Ezek az mRNS-ek biomarkerként szolgálhatnak a betegség korai felismerésében vagy a terápia monitorozásában.
- In vitro diagnosztika: Az mRNS-alapú tesztek gyorsan és pontosan képesek azonosítani a kórokozókat vagy specifikus genetikai variánsokat.
Jövőbeli perspektívák és kutatási irányok

Az mRNS technológia az orvostudomány egyik leggyorsabban fejlődő területe, és a jövőben várhatóan még számos áttörést hoz. A COVID-19 vakcinák sikere csak a kezdet volt, és a kutatók már most is azon dolgoznak, hogy az mRNS potenciálját a lehető legszélesebb körben kihasználják.
Újabb mRNS alapú terápiák
A jövőben számos új mRNS alapú terápia várható:
- Influenza elleni univerzális vakcinák: Az mRNS technológia lehetővé teheti olyan influenza vakcinák kifejlesztését, amelyek egyetlen oltással több törzs ellen is védelmet nyújtanak, vagy akár univerzális védelmet biztosítanak az influenza vírusok ellen.
- Más fertőző betegségek elleni vakcinák: MRNS vakcinák fejlesztése folyik HIV, Zika, Ebola, RSV és más súlyos fertőző betegségek ellen.
- Autoimmun betegségek kezelése: Az mRNS felhasználható lehet olyan fehérjék termelésére, amelyek elnyomják a túlműködő immunválaszt autoimmun betegségek, például sclerosis multiplex vagy rheumatoid arthritis esetén.
- Szív- és érrendszeri betegségek: Az mRNS segíthet a sérült szívizom regenerálásában, vagy olyan fehérjék termelésében, amelyek csökkentik a koleszterinszintet.
- Ritka betegségek: Számos ritka genetikai betegséget okoz egyetlen gén mutációja, amely hiányzó vagy hibás fehérjét eredményez. Az mRNS terápia ideiglenesen pótolhatja ezeket a fehérjéket, enyhítve a tüneteket.
Szállítási rendszerek fejlesztése
Az mRNS stabilitása és hatékony sejtekbe juttatása továbbra is kulcsfontosságú kihívás. A jövőbeli kutatások a lipid nanorészecskék (LNP-k) továbbfejlesztésére fognak koncentrálni, hogy még hatékonyabbak, biztonságosabbak és specifikusabbak legyenek. Célzott LNP-k kifejlesztése, amelyek csak bizonyos sejttípusokba juttatják be az mRNS-t, jelentősen növelné a terápiás potenciált. Emellett más szállítási rendszerek, például polimer alapú nanorészecskék vagy exoszómák vizsgálata is folyik.
Az mRNS módosításának lehetőségei
Az mRNS molekulák módosítása, például pszeudouridin vagy N1-metilpszeudouridin beépítése (mint a COVID-19 vakcinákban), jelentősen javította a transzláció hatékonyságát és csökkentette az immunogenitást. A jövőben további nukleotid módosítások és az mRNS szerkezeti optimalizálása várható, amelyek tovább növelhetik a terápiás mRNS molekulák stabilitását, transzlációs hatékonyságát és biztonságosságát.
Egyénre szabott gyógyászat
Az mRNS technológia kiválóan alkalmas az egyénre szabott gyógyászatra. Különösen a rákterápiában, ahol a betegek tumorai egyedi mutációkat hordoznak, az mRNS vakcinák gyorsan és rugalmasan adaptálhatók az egyes betegek igényeihez. Ez a megközelítés lehetővé teszi a precíziós orvoslás új korszakát, ahol a kezeléseket pontosan a beteg genetikai és molekuláris profiljához igazítják.
Összességében a hírvivő RNS, ez a látszólag egyszerű molekula, a modern orvostudomány egyik legizgalmasabb és legígéretesebb eszköze. A szerkezetének, funkciójának és működésének mélyebb megértése alapvető fontosságú ahhoz, hogy teljes mértékben kiaknázzuk a benne rejlő potenciált, és új utakat nyissunk meg a betegségek megelőzésében és kezelésében.







