
A szervezetünkben zajló biokémiai folyamatok komplex hálózatot alkotnak, melynek minden eleme precízen összehangolt. Ebben a bonyolult rendszerben a lipoproteinek és az általuk szállított anyagok, mint például a koleszterin, központi szerepet játszanak. A „Low Density Lipoprotein-koleszterinaz” kifejezés, bár nem egy standard, tankönyvi enzimnév, arra a kérdéskörre hívja fel a figyelmet, hogy milyen enzimatikus mechanizmusok révén bomlik le, vagy alakul át a Low Density Lipoprotein (LDL) által szállított koleszterin. Valójában nem létezik egyetlen, dedikált „LDL-koleszterinaz” nevű enzim, amely közvetlenül az intakt LDL-részecskéken hatna. Ehelyett számos enzim és transzporter molekula működik együtt a koleszterin metabolizmusában, melyek közül több is érinti az LDL-t, vagy annak koleszterin tartalmát, különösen a sejtekbe való felvétel után. Ennek a cikknek a célja, hogy részletesen feltárja az LDL-koleszterin metabolizmusában részt vevő kulcsfontosságú enzimeket és folyamatokat, megvilágítva azok jelentőségét a szervezet működésében és az egészség megőrzésében.
A koleszterin egy létfontosságú szteroid lipid, amely számos biológiai funkciót tölt be. Sejthártyáink integritásának és fluiditásának fenntartásában elengedhetetlen, előanyaga a szteroid hormonoknak (pl. tesztoszteron, ösztrogén, kortizol), a D-vitaminnak és az epesavaknak. Mivel a koleszterin zsíroldékony, a vérplazmában történő szállítása speciális „csomagolást” igényel. Ezek a csomagolások a lipoproteinek, amelyek egy hidrofób magból (koleszterin-észterek és trigliceridek) és egy hidrofil külső rétegből (foszfolipidek, szabad koleszterin és apolipoproteinek) állnak. Az LDL, vagyis az alacsony sűrűségű lipoprotein, az egyik fő koleszterinszállító részecske a vérben, amely a májból a perifériás szövetekbe juttatja a koleszterint.
Az LDL-részecske szerkezete és funkciója
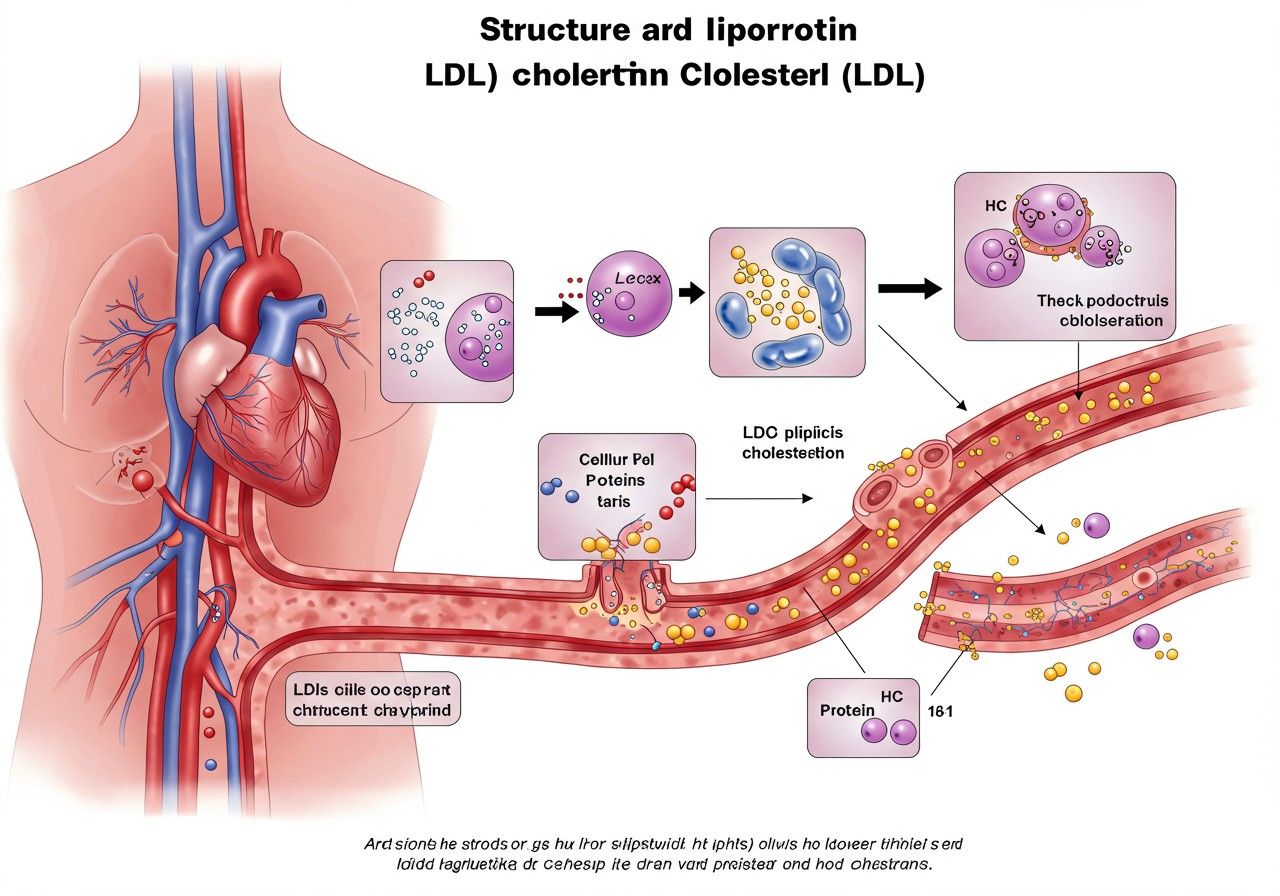
Az LDL-részecske egy gömb alakú struktúra, amelynek magját körülbelül 1500 molekula koleszterin-észter és mintegy 500 molekula triglicerid alkotja. Ezt a hidrofób magot egy foszfolipid monoréteg, szabad koleszterin és egyetlen, nagyméretű fehérje, az apolipoprotein B-100 (apoB-100) veszi körül. Az apoB-100 nemcsak szerkezeti eleme az LDL-nek, hanem kulcsfontosságú ligandumként is szolgál az LDL-receptorok számára, lehetővé téve a részecske felvételét a sejtekbe.
Az LDL elsődleges funkciója a koleszterin szállítása a májból, ahol szintetizálódik vagy a táplálékból szívódik fel, a test sejtjeihez. A sejtek ezt a koleszterint aztán felhasználják membránjaik építéséhez, hormonok szintéziséhez vagy raktározzák. Ez a folyamat nélkülözhetetlen a normális sejtélethez és a szöveti működéshez. Azonban, ha az LDL-koleszterin szintje tartósan magas a vérben, az növeli az érelmeszesedés (atherosclerosis) kockázatát, ami szív- és érrendszeri betegségekhez vezethet.
A koleszterin metabolizmusának kulcsfontosságú enzimei és az LDL kapcsolata
Ahogyan már említettük, a „Low Density Lipoprotein-koleszterinaz” mint egyetlen, specifikus enzim nem létezik. Ehelyett a koleszterin-észterek hidrolízisében és a koleszterin metabolizmusában számos enzim vesz részt, amelyek közül néhány közvetlenül, mások közvetve kapcsolódnak az LDL-hez vagy annak koleszterin tartalmához. Vizsgáljuk meg ezeket részletesebben.
Liszoszomális savas lipáz (LAL) – a legközelebbi analóg
A liszoszomális savas lipáz (LAL) az az enzim, amely a leginkább megfelel a „koleszterinaz” funkciónak az LDL metabolizmusában. Amikor az LDL-részecskék az apoB-100 ligandum révén kötődnek az LDL-receptorokhoz a sejtfelszínen, endocytosis útján bejutnak a sejtbe. Az endoszómákból a lizoszómákba kerülnek, ahol savas pH-n a LAL enzim hidrolizálja az LDL magjában található koleszterin-észtereket és triglicerideket. Ennek eredményeként szabad koleszterin és zsírsavak szabadulnak fel, amelyek aztán kijutnak a lizoszómákból a citoplazmába, hogy a sejt felhasználja őket.
A LAL aktivitásának hiánya vagy csökkent működése súlyos betegségeket okozhat. A legsúlyosabb forma a Wolman-kór, amely csecsemőkorban jelentkezik, és a koleszterin-észterek és trigliceridek felhalmozódásához vezet a májban, lépben és más szervekben, súlyos fejlődési rendellenességeket és korai halált okozva. Egy enyhébb forma a koleszterin-észter tárolási betegség (CESD), amely felnőttkorban is megnyilvánulhat, és hepatomegáliával, májcirrózissal és gyorsított atherosclerosis-szal jár. Ezek a betegségek rávilágítanak a LAL döntő szerepére a koleszterin homeosztázisában és az LDL által szállított koleszterin megfelelő feldolgozásában.
Lipoprotein lipáz (LPL) és hepatikus lipáz (HL)
Bár az lipoprotein lipáz (LPL) és a hepatikus lipáz (HL) elsősorban a trigliceridek hidrolízisében játszanak szerepet, működésük közvetve befolyásolja az LDL kialakulását és metabolizmusát. Az LPL a kapillárisok endothel sejtjeinek felszínén található enzim, amely a kilomikronokban és VLDL-ben (Very Low Density Lipoprotein) lévő triglicerideket hidrolizálja zsírsavakra és glicerinre. Ez a folyamat a VLDL-t VLDL-maradványokká, majd LDL-lé alakítja át. A HL a májban található, és szintén triglicerideket és foszfolipideket hidrolizál a lipoproteinekben, különösen az IDL-ből (Intermediate Density Lipoprotein) LDL-lé történő átalakulásban és a HDL (High Density Lipoprotein) metabolizmusában játszik szerepet.
Ezek az enzimek tehát nem közvetlenül „koleszterinazok”, de a lipoprotein részecskék triglicerid tartalmának csökkentésével hozzájárulnak a részecskék sűrűségének növekedéséhez és végső soron az LDL képződéséhez. Így befolyásolják az LDL koncentrációját és minőségét a keringésben.
Koleszterin-észter transzfer protein (CETP)
A koleszterin-észter transzfer protein (CETP) egy plazmafehérje, amely a koleszterin-észtereket szállítja a HDL-ről a VLDL-re, IDL-re és LDL-re, cserébe trigliceridekért. Ez a folyamat kulcsfontosságú a koleszterin fordított transzportjában (reverse cholesterol transport), de egyben befolyásolja az LDL részecskék összetételét is, növelve azok koleszterin-észter tartalmát és csökkentve a triglicerid tartalmukat. A CETP aktivitásának gátlása potenciális terápiás célpont a HDL-szint növelésére, de klinikai vizsgálatok vegyes eredményeket hoztak az LDL-re gyakorolt hatás és a kardiovaszkuláris események szempontjából.
Lecitin-koleszterin aciltranszferáz (LCAT)
A lecitin-koleszterin aciltranszferáz (LCAT) egy plazmaenzim, amely a HDL metabolizmusában játszik kulcsszerepet. Katalizálja a szabad koleszterin észterifikálását koleszterin-észterré, egy zsírsav átvitelével a lecitinről a koleszterinre. Ez a reakció a HDL részecskékben történik, és a szabad koleszterin hidrofób koleszterin-észterré történő átalakítása lehetővé teszi, hogy a koleszterin a HDL magjába kerüljön, ezzel elősegítve a HDL érését és a koleszterin felvételét a perifériás sejtekből. Bár az LCAT közvetlenül nem hat az LDL-re, a koleszterin dinamikájában betöltött szerepe révén befolyásolja a koleszterin keringését a lipoproteinek között, így közvetetten az LDL-koleszterin szinteket is.
Acil-CoA:koleszterin aciltranszferáz (ACAT)
Az acil-CoA:koleszterin aciltranszteráz (ACAT) egy intracelluláris enzim, amely a szabad koleszterint koleszterin-észterré észterifikálja a sejtekben történő raktározás céljából. Két izoformája ismert: ACAT1 és ACAT2. Az ACAT1 szinte minden sejtben megtalálható, míg az ACAT2 elsősorban a májban és a bélben fejeződik ki. Amikor az LDL által szállított koleszterin bejut a sejtbe és szabad koleszterinként felszabadul a lizoszómákból, az ACAT enzim aktiválódhat, ha a sejt koleszterin szintje magas. Ezáltal a sejt képes raktározni a felesleges koleszterint, megakadályozva annak toxikus felhalmozódását. Az ACAT tehát egy kulcsfontosságú enzim a sejten belüli koleszterin homeosztázis fenntartásában.
HMG-CoA reduktáz
A HMG-CoA reduktáz a koleszterin bioszintézisének sebességmeghatározó enzime. Katalizálja a 3-hidroxi-3-metilglutaril-koenzim A (HMG-CoA) mevalonáttá történő redukcióját, amely a koleszterin szintézis útvonalának korai lépése. Bár ez az enzim nem bontja le a koleszterint, hanem szintetizálja, közvetlenül befolyásolja a máj koleszterin szintjét, és ezáltal az LDL-receptorok expresszióját. Ha a májban alacsony a koleszterin szint, az LDL-receptorok száma megnő a sejtfelszínen, ami több LDL felvételét eredményezi a vérből, csökkentve ezzel a plazma LDL-koleszterin szintjét. A statinok, a leggyakrabban felírt koleszterinszint-csökkentő gyógyszerek, a HMG-CoA reduktáz kompetitív gátlásával fejtik ki hatásukat.
Az LDL-receptor és a koleszterin felvételének szabályozása
Az LDL-receptorok (LDLR) a sejtfelszínen elhelyezkedő glikoproteinek, amelyek specifikusan kötik az apoB-100-at tartalmazó LDL-részecskéket. Ez a kötődés indítja el az endocytosis folyamatát, amelynek során az LDL-részecskék bejutnak a sejtbe. Az LDL-receptorok száma és aktivitása szigorúan szabályozott, elsősorban a sejten belüli koleszterinszint függvényében. Amikor a sejtben alacsony a koleszterin, a szterol szabályozó elem kötőfehérjék (SREBP-k) aktiválódnak, és elősegítik az LDLR gén transzkripcióját, növelve ezzel a receptorok számát a sejtfelszínen. Ezáltal a sejt több LDL-t tud felvenni a vérből, pótolva a koleszterinhiányt.
Ezzel ellentétben, ha a sejten belüli koleszterinszint magas, az SREBP-k inaktiválódnak, és az LDLR gén expressziója csökken. Ezenkívül egy másik fehérje, a PCSK9 (Proprotein Konvertáz Szubtilizin/Kexin Típus 9) is szerepet játszik az LDLR szabályozásában. A PCSK9 az LDLR-hez kötődik, és elősegíti annak lebomlását a lizoszómákban, csökkentve ezzel a sejtfelszínen lévő receptorok számát. Ennek gátlása (pl. PCSK9-gátló gyógyszerekkel) az LDLR szintjének növelésével hatékonyan csökkenti az LDL-koleszterin szintet a vérben.
Az LDL-receptorok működési zavara, mint például a familiáris hypercholesterinaemia (FH) esetén, súlyosan megemelkedett LDL-koleszterinszinthez és korai érelmeszesedéshez vezet.
A koleszterin-észterek és a szabad koleszterin közötti különbség

Fontos megkülönböztetni a szabad koleszterint és a koleszterin-észtereket. A szabad koleszterin egy hidroxil csoporttal rendelkezik a 3-as szénatomon, ami lehetővé teszi, hogy beépüljön a sejtmembránokba, és befolyásolja azok fluiditását. A koleszterin-észterekben ez a hidroxil csoport egy zsírsavval van észterkötésben, ami sokkal hidrofóbbá teszi a molekulát. Ez a forma ideális a koleszterin raktározására és szállítására a lipoproteinek magjában, mivel nem tud beépülni a membránokba. Ahhoz, hogy a koleszterin-észtereket a sejtek felhasználhassák, először hidrolizálni kell szabad koleszterinné, és pontosan ezt a feladatot látja el a lizoszómális savas lipáz (LAL) az LDL által szállított koleszterin esetében.
Az LDL-koleszterin és az atherosclerosis
Az LDL-koleszterin, különösen annak oxidált formája (oxLDL), kulcsszerepet játszik az atherosclerosis, azaz az érelmeszesedés kialakulásában. Amikor az LDL-szint tartósan magas, az LDL-részecskék hajlamosak felhalmozódni az artériák falában, különösen a sérült endothel alatt. Ott oxidatív stressz hatására oxidálódnak, és az oxidált LDL-t a makrofágok „felismerik” és felveszik, ami habsejtek kialakulásához vezet. Ezek a habsejtek az atheromás plakkok magját képezik, amelyek idővel növekednek, szűkítik az ereket, és gyulladásos folyamatokat indítanak el. Végül a plakkok megrepedhetnek, trombózist okozva, ami szívinfarktushoz vagy stroke-hoz vezethet.
Az LDL-koleszterin nem „rossz” önmagában, hiszen létfontosságú a sejtjeink számára. A probléma a magas, tartósan emelkedett szintjével és annak oxidációjával kezdődik, ami az érelmeszesedés fő hajtóereje.
A gyulladásos folyamatok, amelyek az atherosclerosis kialakulásában kulcsszerepet játszanak, szorosan összefüggnek az oxidált LDL jelenlétével. Az oxLDL aktiválja az endothel sejteket, elősegíti a monocyták tapadását és bejutását az érfalba, ahol makrofágokká differenciálódnak. Ezek a makrofágok aztán nagymennyiségű oxLDL-t fagocitálnak, és habsejtekké alakulnak, tovább súlyosbítva a gyulladást és a plakk növekedését. A koleszterin metabolizmusában részt vevő enzimek és transzporterek, mint például a LAL, a CETP, vagy az ACAT, mind befolyásolhatják az LDL-részecskék összetételét, a koleszterin felvételét és raktározását, így közvetve vagy közvetlenül hozzájárulnak az atheroscleroticus folyamatokhoz.
Diagnosztikai jelentőség és célértékek
Az LDL-koleszterin szintjének mérése a lipidprofil részeként alapvető fontosságú a kardiovaszkuláris kockázat felmérésében. A „rossz koleszterin” néven is emlegetett LDL-C célértékei a beteg egyéni kockázati tényezőitől függően változnak. Magas kockázatú egyének, például akiknek már volt szívinfarktusa, stroke-ja, vagy cukorbetegségben szenvednek, sokkal alacsonyabb LDL-C szintre törekednek, mint az alacsony kockázatú populáció. Az elmúlt években egyre nagyobb hangsúlyt kap a non-HDL-koleszterin és az apoB mérése is, mivel ezek pontosabban tükrözik az összes atherogén lipoprotein részecske számát a vérben, mint pusztán az LDL-C.
A non-HDL-koleszterin az összes koleszterin mínusz a HDL-koleszterin, így magában foglalja az LDL-C-n kívül a VLDL-C-t, IDL-C-t és kilomikron-maradvány koleszterint is. Az apoB mérése pedig közvetlenül a koleszterint szállító részecskék számát adja meg, mivel minden atherogén lipoprotein részecske (LDL, VLDL, IDL, Lp(a)) egyetlen apoB molekulát tartalmaz. Ezért az apoB szint emelkedése közvetlenül jelzi az atherogén részecskék megnövekedett számát, függetlenül azok koleszterin tartalmától.
Terápiás stratégiák az LDL-koleszterin szintjének szabályozására
Az LDL-koleszterin szintjének csökkentése az atherosclerosis progressziójának lassításában és a kardiovaszkuláris események kockázatának mérséklésében kulcsfontosságú. Számos terápiás megközelítés létezik, amelyek a koleszterin metabolizmus különböző pontjain avatkoznak be, gyakran az enzimek és transzporterek működését célozva.
Statinok
A statinok a leggyakrabban alkalmazott lipidszint-csökkentő gyógyszerek. Hatásmechanizmusuk a HMG-CoA reduktáz enzim gátlásán alapul, amely a májban a koleszterin bioszintézisének sebességmeghatározó enzime. A koleszterinszintézis csökkenése a májban a sejten belüli koleszterinszint csökkenéséhez vezet, ami kompenzáló mechanizmusként az LDL-receptorok számának növekedését eredményezi a májsejtek felszínén. Ezáltal a máj több LDL-t képes felvenni a vérből, jelentősen csökkentve a plazma LDL-koleszterin szintjét.
Ezetimib
Az ezetimib egy másik koleszterinszint-csökkentő gyógyszer, amely a koleszterin bélből történő felszívódását gátolja. Specifikusan a Niemann-Pick C1-Like 1 (NPC1L1) fehérjét célozza meg, amely a koleszterin transzportereként működik a bélhámsejtekben. Az ezetimib csökkenti a táplálékból és az epéből származó koleszterin felszívódását, ami szintén a máj koleszterinszintjének csökkenéséhez és az LDL-receptorok fokozott expressziójához vezet, hasonlóan a statinokhoz.
PCSK9-gátlók
A PCSK9-gátlók egy újabb generációs lipidszint-csökkentő gyógyszerek, amelyek monoklonális antitestek formájában léteznek (pl. evolocumab, alirocumab). Ezek a gyógyszerek a PCSK9 fehérjéhez kötődnek, és megakadályozzák, hogy az az LDL-receptorokhoz kapcsolódjon és azok lebomlását okozza. A PCSK9 gátlásával az LDL-receptorok tovább maradnak a sejtfelszínen, és folyamatosan képesek felvenni az LDL-t a vérből, ami drámai mértékben csökkenti az LDL-koleszterin szintjét. Különösen hatékonyak azoknál a betegeknél, akik statin intoleranciában szenvednek, vagy familiáris hypercholesterinaemia miatt extrém magas LDL-szinttel rendelkeznek.
Fibrátok
A fibrátok elsősorban a trigliceridszint csökkentésére és a HDL-koleszterinszint növelésére szolgálnak. Hatásmechanizmusuk a PPARα (Peroxiszóma Proliferátor Aktivált Receptor alfa) aktiválásán keresztül valósul meg. Ez a nukleáris receptor szabályozza a lipidmetabolizmusban szerepet játszó gének expresszióját, beleértve az LPL szintézisét és aktivitását is. Az LPL aktivitásának növelésével a fibrátok fokozzák a trigliceridek lebontását a VLDL-ben és kilomikronokban, csökkentve ezzel a trigliceridszintet és közvetve befolyásolva az LDL képződését.
Niacin (B3-vitamin)
A niacin (nikotinsav) nagy dózisban alkalmazva szintén képes csökkenteni az LDL-koleszterin és a trigliceridszintet, miközben növeli a HDL-koleszterin szintjét. Hatásmechanizmusa összetett, és magában foglalja a májban a VLDL szintézisének csökkentését, valamint a HDL katabolizmusának gátlását. Bár hatékony, mellékhatásai, mint például az úgynevezett „niacin flush” (kipirulás, viszketés), korlátozzák széleskörű alkalmazását.
Életmódbeli változtatások
A gyógyszeres terápia mellett az egészséges életmód alapvető fontosságú az LDL-koleszterin szintjének szabályozásában. Ide tartozik a telített és transzzsírsavakban szegény, rostban gazdag étrend, a rendszeres testmozgás, az optimális testsúly fenntartása és a dohányzás mellőzése. Ezek a változtatások befolyásolják a koleszterin felszívódását, szintézisét és metabolizmusát, hozzájárulva a lipidprofil javulásához.
Jövőbeli kutatási irányok és potenciális terápiás célpontok

A koleszterin metabolizmusának bonyolult hálózatának mélyebb megértése folyamatosan új terápiás célpontokat azonosít. A „Low Density Lipoprotein-koleszterinaz” fogalmának mélyére ásva, és az LDL-koleszterin metabolizmusában részt vevő enzimek és mechanizmusok tanulmányozásával számos potenciális terület tárul fel a jövőbeni gyógyszerfejlesztés számára.
ANGPTL3 és ANGPTL4 gátlása
Az ANGPTL3 (Angiopoietin-Like 3) és ANGPTL4 (Angiopoietin-Like 4) fehérjék a lipoprotein lipáz (LPL) és a hepatikus lipáz (HL) endogén gátlói. Ezeknek a fehérjéknek a gátlása (pl. monoklonális antitestekkel) az LPL és HL aktivitásának növeléséhez vezet, ami a trigliceridszint csökkenését és a HDL-szint növekedését eredményezi. Az ANGPTL3 gátlása az LDL-koleszterin szintjét is csökkenti, mivel befolyásolja a VLDL metabolizmusát.
Adenozin trifoszfát-citrát liáz (ACL) gátlása
Az adenozin trifoszfát-citrát liáz (ACL) egy enzim, amely a citrátot acetil-CoA-vá alakítja a citoplazmában, amely azután a koleszterin és a zsírsavak szintéziséhez szükséges prekurzor. Az ACL gátlása (pl. bempedoesavval) csökkenti a koleszterinszintézist a májban, hasonlóan a statinokhoz, és így csökkenti az LDL-koleszterin szintjét. Ez egy orális gyógyszer, amely alternatívát kínálhat a statin intoleranciában szenvedők számára.
MikroRNS-ek és génterápia
A mikroRNS-ek (miRNA-k) olyan rövid, nem kódoló RNS molekulák, amelyek poszt-transzkripciós szinten szabályozzák a génexpressziót. Számos miRNA részt vesz a lipid metabolizmus szabályozásában, és potenciális terápiás célpontokat jelenthetnek. Például a miR-33 gátlása növelheti a HDL-szintet és csökkentheti az LDL-szintet. A génterápiás megközelítések, különösen a familiáris hypercholesterinaemia súlyos formáiban, szintén ígéretesek lehetnek az LDL-receptorok expressziójának helyreállításában vagy a PCSK9 gátlásában.
Egyénre szabott orvoslás
A genetikai variációk és az egyéni metabolikus profilok egyre mélyebb megértése lehetővé teszi a jövőben az egyénre szabott orvoslást. Ez azt jelenti, hogy a betegek számára a legmegfelelőbb lipidszint-csökkentő terápiát választhatjuk ki, figyelembe véve genetikai hajlamaikat, egyéb betegségeiket és a gyógyszerekre adott egyéni válaszaikat. Ez optimalizálhatja a kezelés hatékonyságát és minimalizálhatja a mellékhatásokat.
Az LDL-koleszterin metabolizmusának komplexitása rávilágít arra, hogy a „Low Density Lipoprotein-koleszterinaz” fogalma mögött egy egész sor enzim, transzporter és szabályozó mechanizmus rejtőzik. A lizoszomális savas lipáz (LAL) a legközvetlenebb enzim, amely a sejtekbe felvett LDL koleszterin-észtereket hidrolizálja. Azonban az LPL, HL, CETP, LCAT, ACAT és HMG-CoA reduktáz is mind kulcsszerepet játszanak a koleszterin homeosztázisában és az LDL szintjének szabályozásában. Ezen folyamatok alapos megértése elengedhetetlen a kardiovaszkuláris betegségek megelőzéséhez és kezeléséhez, valamint új, innovatív terápiás stratégiák kidolgozásához.







