
A Földön zajló élet alapja a fotoszintézis, az a lenyűgöző biokémiai folyamat, amely során a növények, algák és bizonyos baktériumok a napfény energiáját kémiai energiává alakítják. Ennek a csodálatos transzformációnak a kulcsszereplője egy zöld pigment, a klorofill a. Ez a molekula nem csupán a növényvilág jellegzetes színéért felelős, hanem a fényenergia befogásának és továbbításának elsődleges eszköze is, nélküle a földi élet, ahogyan ismerjük, nem létezhetne.
A klorofill a egy univerzális pigment, amely szinte minden fotoszintetikus eukariótában (növények, algák) és cianobaktériumban megtalálható. Kémiai felépítése rendkívül specializált, lehetővé téve számára a fénykvantumok rendkívül hatékony elnyelését és az így nyert energia célzott átadását a fotoszintetikus reakcióközpontokba. Megértése elengedhetetlen a fotoszintézis komplex mechanizmusának feltárásához, valamint az ökológiai rendszerek működésének és az élet energetikai alapjainak mélyebb megismeréséhez.
Ez a cikk részletesen bemutatja a klorofill a szerkezetét, elmélyedve a molekula kémiai felépítésének finomságaiban, amelyek lehetővé teszik egyedülálló funkcióját. Vizsgáljuk a klorofill a szerepét a fotoszintézisben, kitérve a fénygyűjtő komplexekben és a reakcióközpontokban betöltött feladataira, valamint az energiaátadás alapvető mechanizmusaira. Végül elemezzük a klorofill a abszorpciós spektrumát, magyarázatot adva arra, hogy mely hullámhosszú fényt nyeli el, és miért bocsátja vissza a zöld fényt, ami a növények jellegzetes színét adja.
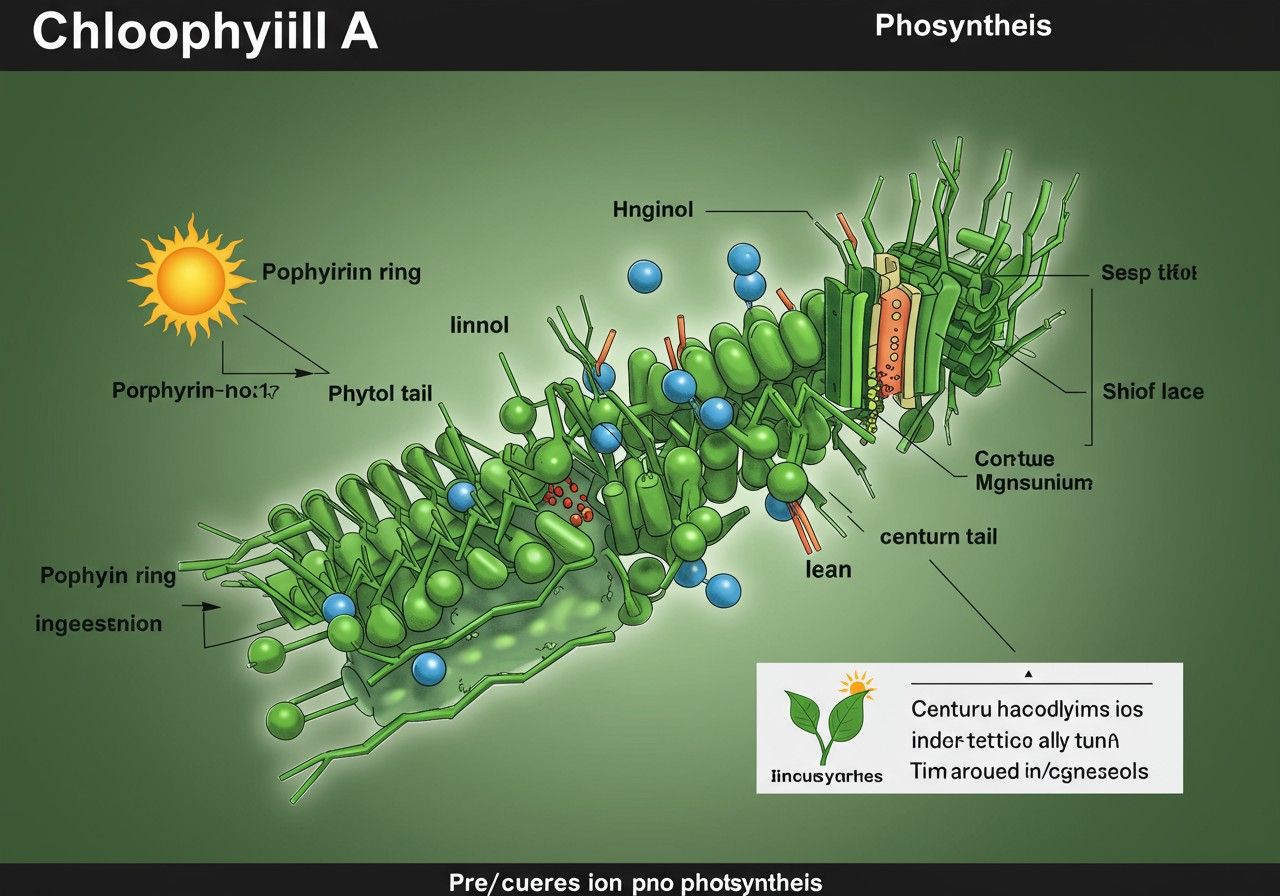
A klorofill a kémiai szerkezete: egy mesterien megtervezett molekula
A klorofill a egy lenyűgözően komplex, ám rendkívül funkcionális szerves molekula, amelynek kémiai szerkezete kulcsfontosságú a fényenergia hatékony befogásában. Alapvetően két fő részből áll: egy sík, gyűrűs struktúrából, az úgynevezett porfirin gyűrűből, és egy hosszú, hidrofób szénhidrogén láncból, a fitol farokból.
A porfirin gyűrű a klorofill a funkcionális magja, amely négy pirrolgyűrűből áll, ezek metin hidak (–CH=) kapcsolják össze egy nagyobb, konjugált rendszerré. Ennek a makrociklusos szerkezetnek a közepén egy magnézium (Mg2+) ion található, amely kovalens és koordinációs kötésekkel kapcsolódik a négy pirrolgyűrű nitrogénatomjaihoz. Ez a magnéziumion nélkülözhetetlen a klorofill fényelnyelő képességéhez, mivel befolyásolja a gyűrű elektronrendszerének energiáját, és így a molekula abszorpciós tulajdonságait.
A porfirin gyűrű körüli oldalláncok nem csupán a molekula stabilitásáért felelősek, hanem finomhangolják annak fényelnyelő képességét is. A klorofill a esetében a 7-es pozícióban egy metilcsoport (-CH₃) található, ami megkülönbözteti a klorofill b-től, ahol ezen a helyen egy formilcsoport (-CHO) helyezkedik el. Ez a különbség, bár aprónak tűnhet, jelentősen befolyásolja a két pigment abszorpciós spektrumát, lehetővé téve számukra, hogy a fény különböző hullámhosszait nyeljék el, optimalizálva a fénygyűjtést.
A porfirin gyűrűhöz kovalensen kapcsolódik a fitol farok, egy 20 szénatomos, elágazó láncú alkohol. Ez a hosszú, hidrofób rész kulcsfontosságú a klorofill a membránokba való beágyazódásában. A fitol farok biztosítja, hogy a klorofill molekula stabilan rögzüljön a tilakoid membrán lipid kettős rétegében, ahol a fotoszintézis folyamatai zajlanak. Enélkül a membránhoz kötődés nélkül a klorofill nem tudná hatékonyan elvégezni a fénygyűjtő és energiaátadó funkcióját.
A molekula konjugált kettős kötésrendszere, amely a porfirin gyűrűben terjed ki, teszi lehetővé a fény elnyelését. Az elektronok ezen a kiterjedt rendszeren belül delokalizáltak, ami azt jelenti, hogy nem egyetlen atomhoz vagy kötéshez tartoznak, hanem az egész gyűrűn eloszlanak. Amikor egy foton, azaz egy fénykvantum energiája eléri a klorofill a molekulát, az elektronok a stabil alapállapotból egy magasabb energiájú, gerjesztett állapotba ugranak. Ez a gerjesztett állapot azonban instabil, és az energia azonnal továbbadódik a fotoszintetikus rendszeren belül.
A klorofill a porfirin gyűrűjének középpontjában elhelyezkedő magnéziumion, valamint a kiterjedt konjugált rendszer együttesen biztosítja a molekula kivételes fényelnyelő képességét, ami a fotoszintézis alapját képezi.
A klorofill a szerkezeti sokszínűsége a természetben is megfigyelhető. Bár a klorofill a a legelterjedtebb forma, léteznek más klorofill típusok is, mint például a klorofill b, c, d, és f, amelyek mindegyike apró kémiai különbségekkel rendelkezik az oldalláncaikban, ami eltérő abszorpciós spektrumot és ezáltal eltérő fényhasznosítási stratégiát eredményez. Ezek a molekuláris finomhangolások teszik lehetővé, hogy a különböző fotoszintetikus szervezetek a környezetükben rendelkezésre álló fényforrások széles spektrumát a lehető leghatékonyabban hasznosítsák.
A klorofill a szerkezete tehát egy tökéletes példa arra, hogyan illeszkedik a molekuláris szintű felépítés a biológiai funkcióhoz. A porfirin gyűrű, a magnéziumion és a fitol farok harmonikus együttműködése biztosítja a fényenergia befogásának és továbbításának precíz mechanizmusát, amely nélkülözhetetlen a földi élet fenntartásához.
A klorofill a szerepe a fotoszintézisben: az élet motorja
A klorofill a nem csupán egy pigment, hanem a fotoszintézis, azaz a fényenergia kémiai energiává alakításának központi molekulája. Szerepe kettős és egymással szorosan összefüggő: egyrészt a fénygyűjtő komplexek (antenna komplexek) fő alkotóelemeként gyűjti és továbbítja a fényenergiát, másrészt a reakcióközpontokban (fotoszisztemák) elindítja a fotokémiai reakciókat, amelyek az elektrontranszport lánc alapját képezik.
A fénygyűjtő komplexek és az energiaátadás
A legtöbb klorofill a molekula nem közvetlenül vesz részt a kémiai reakciókban, hanem antenna pigmentként funkcionál. Ezek a molekulák, más pigmentekkel (pl. klorofill b, karotinoidok) együtt, szorosan szervezett komplexekbe, az úgynevezett fénygyűjtő komplexekbe (LHC – Light-Harvesting Complex) rendeződnek a tilakoid membránban. Az LHC feladata a beérkező fényenergia maximalizálása és hatékony továbbítása a reakcióközpontok felé.
Amikor egy foton eléri a fénygyűjtő komplexben lévő klorofill a vagy más pigment molekulát, az elnyelt energia gerjeszti az elektronjait. Ez a gerjesztési energia nem vész el hő vagy fluoreszcencia formájában, hanem rendkívül gyorsan és hatékonyan, rezonancia energia transzfer (FRET – Förster Resonance Energy Transfer) mechanizmusával továbbítódik a szomszédos pigment molekulákra. Ez a folyamat egy „energia tölcsérként” működik, ahol az energia a magasabb energiájú (rövidebb hullámhosszú fényt elnyelő) pigmentektől a mélyebb energiájú (hosszabb hullámhosszú fényt elnyelő) pigmentek felé áramlik, amíg el nem éri a reakcióközpontot.
A rezonancia energia transzfer egy kulcsfontosságú mechanizmus, amely lehetővé teszi a fotoszintetikus rendszer számára, hogy a beérkező fény széles spektrumát hasznosítsa, és az energiát veszteségmentesen, nanomásodpercek alatt eljuttassa a célállomásra. A klorofill a mint elsődleges antenna pigment biztosítja ennek a „tölcsérnek” a legmélyebb pontjait, ahonnan az energia már közvetlenül a reakcióközpontba kerül.
A reakcióközpontok és a fotokémiai átalakulás
A fénygyűjtő komplexek által gyűjtött energia végül a reakcióközpontokba jut, ahol a klorofill a molekulák egy speciális párja, az úgynevezett primer donor, végzi el a fotokémiai átalakulást. Két fő típusú reakcióközpont létezik a növények és algák tilakoid membránjában:
- Fotoszisztema II (PSII): Ennek reakcióközpontja a P680 nevű klorofill a dimer, amely 680 nm hullámhosszú fényt nyel el a leghatékonyabban. A P680 gerjesztése után lead egy elektront egy akceptor molekulának, és maga oxidálódik (P680+ lesz belőle). Az elvesztett elektront a víz fotolíziséből (vízbontás) nyeri vissza, ami során oxigén (O₂) szabadul fel.
- Fotoszisztema I (PSI): Ennek reakcióközpontja a P700 nevű klorofill a dimer, amely 700 nm hullámhosszú fényt nyel el a leghatékonyabban. A P700 gerjesztése után szintén lead egy elektront egy akceptor molekulának, és oxidálódik (P700+ lesz belőle). Az elvesztett elektront a PSII-ből érkező elektrontranszport lánc végéről kapja vissza.
A reakcióközpontokban zajló elektronátadás az első lépés a fényenergia kémiai energiává alakításában. A gerjesztett klorofill a molekulából kilépő elektronok egy sor redoxreakción keresztül, az elektrontranszport láncon haladnak végig. Ez a lánc különböző fehérjekomplexekből és elektronhordozó molekulákból áll, amelyek a tilakoid membránban helyezkednek el.
A klorofill a a fényenergia kémiai energiává alakításának kulcsfontosságú szereplője, amely a napfényt hasznosítható formába önti, lehetővé téve a szerves anyagok szintézisét és az élet fenntartását.
Az elektrontranszport láncban az elektronok mozgása során felszabaduló energia felhasználódik protonok (H+) pumpálására a tilakoid lumenbe. Ez a proton gradiens, azaz a membrán két oldala közötti koncentrációkülönbség, hajtja az ATP-szintáz enzimet, amely ATP-t (adenozin-trifoszfátot) termel. Az ATP az univerzális energiatároló valuta a sejtekben. Ugyanakkor az elektrontranszport lánc végén a P700-ból kilépő elektronok a NADP+ redukciójához vezetnek NADPH-vá, amely egy erős redukáló ágens, és a szén-dioxid fixálásához szükséges.
Az ATP és NADPH molekulák a fotoszintézis ún. fényfüggő szakaszában termelődnek, és ezek az energiahordozók hajtják a fényfüggetlen szakaszban (Calvin-ciklus) zajló szén-dioxid redukciót és a szénhidrátok szintézisét. A klorofill a tehát közvetlenül indítja el azokat a folyamatokat, amelyek a légköri szén-dioxidot szerves anyaggá alakítják, ezzel biztosítva a földi táplálékláncok alapját.
Összefoglalva, a klorofill a szerepe a fotoszintézisben elengedhetetlen. Mint antenna pigment, maximalizálja a fénygyűjtést, majd mint a reakcióközpontok primer donora, elindítja az elektrontranszport láncot, amely az ATP és NADPH termeléséhez vezet. Ez a komplex, de rendkívül hatékony rendszer teszi lehetővé a napenergia befogását és átalakítását, ami az ökoszisztémák működésének és az élet fenntartásának alapja.
A klorofill a abszorpciós spektruma: miért zöld a növény?
A klorofill a abszorpciós spektruma az a grafikon, amely megmutatja, hogy a molekula milyen hatékonysággal nyeli el a különböző hullámhosszú fénysugarakat. Ez a spektrum nem csupán tudományos érdekesség, hanem alapvetően magyarázza, miért látjuk a növényeket zöldnek, és hogyan optimalizálják a fényenergia hasznosítását.
A spektrum jellegzetességei és magyarázatuk
A klorofill a abszorpciós spektruma két fő elnyelési csúcsot mutat a látható fény tartományában:
- Kék-ibolya régió: Egy erős abszorpciós csúcs található a 430-470 nm közötti hullámhosszon, a kék és ibolya fény tartományában.
- Vörös régió: Egy másik, de valamivel gyengébb abszorpciós csúcs figyelhető meg a 660-680 nm közötti hullámhosszon, a vörös fény tartományában.
Ezek a csúcsok azt jelzik, hogy a klorofill a rendkívül hatékonyan nyeli el a kék és a vörös fényt. A klorofill a molekula kiterjedt konjugált kettős kötésrendszere (a porfirin gyűrűben) felelős ezekért az abszorpciós tulajdonságokért. Amikor egy foton energiája megegyezik a molekula elektronjainak egy alacsonyabb energiaszintről egy magasabb energiaszintre történő gerjesztéséhez szükséges energiával, a fény elnyelődik. A kék fény nagyobb energiájú, a vörös fény alacsonyabb energiájú fotonokból áll, és a klorofill a elektronrendszere pontosan ezeket az energiakülönbségeket tudja áthidalni.
A spektrum legszembetűnőbb jellemzője azonban az, hogy a klorofill a nagyon gyengén nyeli el a zöld fényt (kb. 500-600 nm hullámhosszon). Ehelyett a zöld fény nagy részét visszaveri vagy átereszti. Ez az oka annak, hogy az emberi szem a növényeket zöldnek látja: a zöld fény az, ami eljut a szemünkbe, miután a növény elnyelte a spektrum többi részét a fotoszintézishez.
A klorofill a abszorpciós spektruma egyértelműen mutatja, hogy a növények miért zöldek: a kék és vörös fényt elnyelik a fotoszintézishez, míg a zöld fényt visszaverik, ami a jellegzetes színt adja.
Klorofill a és b közötti különbségek
Fontos megkülönböztetni a klorofill a abszorpciós spektrumát a klorofill b spektrumától. A klorofill b, amely szintén általánosan megtalálható a zöld növényekben és algákban, kiegészítő pigmentként funkcionál, segítve a klorofill a-t a fénygyűjtésben. A klorofill b abszorpciós csúcsai kissé eltolódottak a klorofill a-hoz képest:
| Pigment | Fő abszorpciós csúcsok (nm) | Megjegyzés |
|---|---|---|
| Klorofill a | ~430-470 (kék), ~660-680 (vörös) | Primer fotoszintetikus pigment |
| Klorofill b | ~450-490 (kék), ~640-660 (narancs/vörös) | Kiegészítő pigment, szélesíti az elnyelt spektrumot |
Ez az apró különbség az abszorpciós csúcsokban, mely a klorofill b esetében a 7-es pozícióban lévő formilcsoportnak köszönhető, lehetővé teszi, hogy a klorofill b a fény olyan hullámhosszait is elnyelje, amelyeket a klorofill a kevésbé hatékonyan hasznosít. Az elnyelt energiát aztán továbbítja a klorofill a-nak a fénygyűjtő komplexeken keresztül, így a fotoszintetikus rendszer a napfény szélesebb spektrumát tudja hasznosítani.
A klorofill a abszorpciójának mérése
A klorofill a abszorpciós spektrumát általában spektrofotometriával mérik. Ez a technika magában foglalja a fény áteresztésének vagy elnyelésének mérését egy mintán különböző hullámhosszakon. A klorofillt általában oldószerbe (pl. aceton, etanol) extrahálják a növényi szövetekből, majd az oldat abszorbanciáját mérik. Az így kapott spektrum pontosan feltárja a pigment fényelnyelő tulajdonságait.
A klorofill a abszorpciós spektrumának ismerete alapvető fontosságú a fotoszintézis kutatásában. Segít megérteni, hogyan alkalmazkodnak a növények a különböző fényviszonyokhoz, hogyan optimalizálják a fényhasznosítást, és milyen fényforrásokra van szükségük a hatékony növekedéshez. Ezen ismeretek felhasználhatók a mezőgazdaságban, a hidroponikában és a mesterséges megvilágítású növénytermesztésben is az optimális fényviszonyok megteremtésére.
A klorofill a abszorpciós spektruma tehát nem csak egy fizikai jelenség, hanem a biológiai adaptáció és optimalizáció ragyogó példája, amely a napfény energiáját az élet fenntartásához szükséges kémiai energiává alakítja.
A fotoszintézis komplex gépezete: klorofill a a rendszerben

A klorofill a molekula, bár önmagában is lenyűgöző, csak egy része egy sokkal nagyobb és bonyolultabb rendszernek: a fotoszintetikus apparátusnak. Ennek a gépezetnek a működése a növények tilakoid membránjában zajlik, ahol a klorofill a és más pigmentek, fehérjék, enzimek és elektronhordozók precízen összehangolt együttműködése teszi lehetővé a fényenergia hatékony átalakítását.
A tilakoid membrán és a fotoszisztemák elhelyezkedése
A fotoszintézis fényfüggő reakciói a kloroplasztiszok belső membránrendszerében, a tilakoid membránban zajlanak. Ez a membránrendszer lapos, zsák alakú struktúrákat, a tilakoidokat alkotja, amelyek gyakran egymásra rétegződve, ún. gránumokat formálnak. A tilakoid membrán a klorofill a, klorofill b, karotinoidok, valamint a fénygyűjtő komplexek és reakcióközpontok (Fotoszisztema I és II) fehérjéinek otthona.
A Fotoszisztema II (PSII) és a Fotoszisztema I (PSI) a tilakoid membránba ágyazott nagy fehérje-pigment komplexek. Bár mindkettő tartalmaz klorofill a-t a reakcióközpontjában, elhelyezkedésük és funkciójuk kissé eltérő. A PSII jellemzően a gránumok egymásra rakott részein (grana lamellák) található, míg a PSI inkább a gránumokat összekötő stroma lamellákban, valamint a gránumok szélein helyezkedik el. Ez a térbeli elválasztás optimalizálja a fényhasznosítást és az elektrontranszport lánc működését.
Kiegészítő pigmentek és a fénygyűjtés optimalizálása
Bár a klorofill a a fő fotoszintetikus pigment, a fénygyűjtő komplexekben számos más pigment is jelen van, amelyek kiegészítő szerepet töltenek be. Ezek közé tartozik a klorofill b és a karotinoidok (pl. béta-karotin, lutein). Ezek a kiegészítő pigmentek eltérő abszorpciós spektrummal rendelkeznek, mint a klorofill a, lehetővé téve a növény számára, hogy a látható fény szélesebb tartományát hasznosítsa.
- Klorofill b: Ahogy korábban említettük, a klorofill b abszorpciós csúcsai kissé eltolódottak a klorofill a-hoz képest, különösen a kék-zöld és a narancs-vörös régióban. Ezáltal olyan fényhullámhosszakat is elnyel, amelyeket a klorofill a kevésbé hatékonyan hasznosítana.
- Karotinoidok: Ezek a sárga, narancs vagy vörös pigmentek elsősorban a kék és zöld fény tartományában nyelnek el. Két fő funkciójuk van: egyrészt kiegészítő fénygyűjtőként továbbítják az energiát a klorofill a-nak, másrészt, ami talán még fontosabb, fotoprotektív szerepet töltenek be. Elnyelik a felesleges fényenergiát, amely károsíthatná a klorofill molekulákat (pl. reaktív oxigénfajták képződésével), és hővé alakítva azt elvezetik a rendszerből.
Ez a pigmentek sokfélesége biztosítja, hogy a növény a lehető legtöbb beérkező fényenergiát képes legyen befogni és hasznosítani, még változó fényviszonyok között is. Az elnyelt energia a rezonancia energia transzfer mechanizmusával jut el az összes kiegészítő pigmentből a klorofill a-ba, majd onnan a reakcióközpontokba.
Az elektrontranszport lánc és a protongradiens kialakulása
A reakcióközpontokban (PSII és PSI) elinduló elektronok egy szervezett útvonalon, az elektrontranszport láncon haladnak végig. Ez a lánc több fehérjekomplexből áll, amelyek a tilakoid membránba ágyazódnak:
- A PSII-ből kilépő elektronok először a plasztoquinonra (PQ) kerülnek, amely mozgékony elektronhordozóként funkcionál.
- A PQ redukált formája (PQH₂) továbbítja az elektronokat a citokróm b₆f komplexnek. Ez a komplex egyúttal protonokat (H+) pumpál a tilakoid lumenbe a sztromából, hozzájárulva a protongradiens kialakulásához.
- A citokróm b₆f komplexből az elektronok a plaszto-cianinra (PC) kerülnek, amely egy vízoldékony fehérje a tilakoid lumenben, és továbbítja az elektronokat a PSI-nek.
- A PSI reakcióközpontja (P700) is elnyel fényt és gerjesztődik, majd leadja elektronjait a ferredoxinra (Fd). A P700+ az elektronokat a PC-től kapja vissza.
- A ferredoxin az elektronokat a ferredoxin-NADP+-reduktáz (FNR) enzimnek adja át, amely a NADP+-t NADPH-vá redukálja. A NADPH a Calvin-ciklus redukciós lépéseihez szükséges.
Az elektrontranszport lánc során a tilakoid lumenben felhalmozódó protonok (a vízbontásból és a citokróm b₆f komplex protonpumpálásából származóan) egy protongradienst hoznak létre a tilakoid membrán két oldala között. Ez a gradiens egy elektrokémiai potenciálkülönbséget jelent, amely tárolt energiát képvisel. Amikor a protonok visszajutnak a sztromába egy speciális csatornán, az ATP-szintázon keresztül, az ATP-szintáz ezt az energiát felhasználja ATP szintézisére ADP-ből és anorganikus foszfátból.
Ez a folyamat, a fotofoszforiláció, a fényenergia kémiai energiává (ATP és NADPH) való végső átalakítását jelenti, amelyet a növény aztán a szén-dioxid fixálásához és a szerves vegyületek szintéziséhez használ fel a Calvin-ciklusban. A klorofill a tehát nem csupán elnyeli a fényt, hanem a teljes fotoszintetikus gépezet indítópontja és hajtóereje, amely az életet lehetővé tevő energetikai alapokat biztosítja a Földön.
A klorofill a bioszintézise és lebomlása: egy dinamikus körforgás
A klorofill a nem egy statikus molekula a növényi sejtekben; folyamatosan szintetizálódik és bomlik le, különösen a növekedés és az öregedés során. Ez a dinamikus körforgás biztosítja a fotoszintetikus apparátus optimális működését és a növények alkalmazkodását a változó környezeti feltételekhez.
A klorofill a bioszintézise: a fény szerepe
A klorofill a szintézise egy komplex, több lépésből álló biokémiai útvonal, amelynek kiinduló anyaga a glutamát, egy aminosav. Az útvonal során számos intermedier képződik, amelyek közül a legfontosabb a protoporfirin IX. Ebbe a molekulába épül be aztán a magnézium (Mg2+) ion, létrehozva a magnézium-protoporfirin IX-et.
A bioszintézis egyik legkritikusabb lépése a protochlorophyllide redukciója chlorophyllide a-vá. Ez a lépés a legtöbb növényben fényfüggő, amit a protochlorophyllide oxidoreduktáz (POR) enzim katalizál. Ez az enzim csak fény jelenlétében aktív. Ez az oka annak, hogy a sötétben nevelt növények (etiolált palánták) sárgásfehérek (fehérjék, karotinoidok vannak bennük, de klorofill nincs), és csak fény hatására válnak zölddé. A fény stimulálja a klorofill szintézisét, lehetővé téve a fotoszintetikus apparátus kialakulását.
A chlorophyllide a-hoz kapcsolódik ezután a fitol farok, amelyet a chlorophyll synthase enzim katalizál. Ezzel a lépéssel alakul ki a teljes értékű klorofill a molekula. A bioszintézis folyamata szigorúan szabályozott, és számos gén és enzim összehangolt működését igényli. A fény intenzitása, a hőmérséklet és a tápanyagellátottság mind befolyásolják a klorofill termelődését.
A klorofill a lebomlása: az őszi lombszíneződés
A klorofill a lebomlása ugyanolyan fontos biológiai folyamat, mint a szintézise. Ennek leglátványosabb megnyilvánulása az őszi lombszíneződés. A vegetációs időszak végén, a hidegebb hőmérséklet, a rövidebb nappalok és az intenzívebb fény hatására a növények leállítják a klorofill termelését, és megkezdik a már meglévő klorofill molekulák lebontását.
A lebomlási folyamat a klorofilláz enzim működésével kezdődik, amely leválasztja a fitol farkat a porfirin gyűrűről. Ezután a porfirin gyűrű is több lépésben bomlik le, színezékmentes klorofill katabolitokká (NCCs – Non-fluorescent Chlorophyll Catabolites) alakulva. Ezek a termékek gyakran színtelenek vagy halványsárgák, és a vakuólumban tárolódnak, vagy tovább bomlanak.
Az őszi lombszíneződés nem csupán esztétikai jelenség, hanem egy rendkívül fontos biológiai folyamat, amely során a növények hatékonyan újrahasznosítják a klorofill molekulák értékes alkotóelemeit, mielőtt elveszítenék leveleiket.
A klorofill lebomlásának célja az, hogy a növény újrahasznosítsa a klorofill molekulákban található értékes nitrogént és magnéziumot. Ezek az elemek visszaszállítódnak a növény törzsébe és gyökereibe, ahol tárolódnak, és a következő tavaszi növekedéshez felhasználhatók. A klorofill lebomlása után előtérbe kerülnek a már korábban is jelen lévő, de a zöld klorofill által elfedett karotinoidok (sárga és narancssárga színek), valamint a hideg hatására szintetizálódó antociánok (vörös és lila színek), amelyek együttesen adják az őszi erdők színpompás látványát.
Ez a szintézis-lebomlás ciklus nem csupán az őszi jelenséghez kapcsolódik. A klorofill folyamatosan bomlik le és szintetizálódik a fotoszintetikusan aktív levelekben is, bár sokkal lassabb ütemben. Ez a dinamika lehetővé teszi a növények számára, hogy reagáljanak a stresszre, például a túlzott fényre vagy a tápanyaghiányra, és fenntartsák a fotoszintetikus apparátus optimális állapotát.
A klorofill a bioszintézisének és lebomlásának megértése alapvető fontosságú a növényélettanban és a mezőgazdaságban. Az ezen folyamatokba való beavatkozás lehetőséget kínál a növények növekedésének és terméshozamának optimalizálására, valamint a környezeti stresszre való ellenálló képességük javítására.
Klorofill a a különböző fotoszintetikus szervezetekben: az evolúció sokszínűsége
Bár a klorofill a a legelterjedtebb és legfontosabb fotoszintetikus pigment, jelenléte és szerepe némileg eltérő lehet a különböző fotoszintetikus szervezetekben. Ez a sokszínűség az evolúciós adaptációkat tükrözi, amelyek lehetővé teszik ezen élőlények számára, hogy a legkülönfélébb környezeti feltételek között is sikeresen végezzék a fotoszintézist.
Növények és zöld algák
A szárazföldi növények és a zöld algák a klorofill a mellett jelentős mennyiségű klorofill b-t is tartalmaznak. Ez a két pigment együttműködve szélesíti a fotoszintézishez hasznosítható fény spektrumát. A klorofill b a kék és narancssárga fény elnyelésével kiegészíti a klorofill a abszorpcióját, és az energiát továbbítja a klorofill a-nak a reakcióközpontokba. Emellett karotinoidok is jelen vannak, amelyek fotoprotektív szerepet töltenek be.
Ez a pigmentösszetétel a zöld növények és algák evolúciós sikereinek egyik alapja, lehetővé téve számukra, hogy hatékonyan hasznosítsák a napfényt a szárazföldi és sekélyvízi környezetekben, ahol a teljes látható spektrum elérhető.
Cianobaktériumok (kék-zöld algák)
A cianobaktériumok, amelyek az oxigéntermelő fotoszintézis evolúciós ősei, szintén tartalmaznak klorofill a-t mint primer fotoszintetikus pigmentet. A klorofill b helyett azonban gyakran fikobiliproteineket (fikocianin, fikoeritrin) használnak kiegészítő pigmentként. Ezek a fehérjék rendkívül hatékonyan nyelik el a zöld, sárga és narancssárga fényt, amely mélyebben hatol be a vízbe, mint a kék vagy vörös fény. A fikobiliproteinek az energiát a fikobiliszómáknak nevezett komplexekben gyűjtik, majd továbbítják a klorofill a-nak.
Ez az adaptáció lehetővé teszi a cianobaktériumok számára, hogy sikeresen kolonizálják a vízi környezeteket, beleértve a mélyebb vizeket és az árnyékosabb területeket is, ahol a fényspektrum már eltolódott a zöld és sárga felé.
Vörös algák
A vörös algák, hasonlóan a cianobaktériumokhoz, klorofill a-t tartalmaznak, és kiegészítő pigmentként szintén fikobiliproteineket (különösen fikoeritrint) használnak. A fikoeritrin a zöld és kék fényt nyeli el, ami a vörös algáknak a jellegzetes vörös színét adja, és lehetővé teszi számukra, hogy a tenger mélyebb, kékebb vizeiben is fotoszintetizáljanak, ahová a zöld és sárga fény már nem jut el.
A klorofill a univerzális jelenléte a fotoszintetikus organizmusokban, kiegészítve a különböző segédpigmentekkel, az élet alkalmazkodóképességének és a fényenergia hasznosításának evolúciós sokszínűségének tanúbizonysága.
Barna algák és kovamoszatok
A barna algák és a kovamoszatok (diatómák) szintén tartalmaznak klorofill a-t. Kiegészítő pigmentként azonban klorofill c-t és a fukoxantint, egy karotinoidot használnak. A fukoxantin a zöld és kék-zöld fényt nyeli el, ami a barna algáknak a jellegzetes barnás-olajzöld színét adja. Ez a pigmentösszetétel lehetővé teszi számukra, hogy hatékonyan végezzék a fotoszintézist a sekélyebb, de gyakran zavarosabb tengeri környezetekben.
Anoxigén fotoszintetikus baktériumok
Léteznek olyan fotoszintetikus baktériumok is, amelyek nem termelnek oxigént (ún. anoxigén fotoszintézis), és nem tartalmaznak klorofill a-t. Ehelyett speciális pigmenteket, az úgynevezett baktérioklorofillokat (pl. baktérioklorofill a, b, c, d, e, g) használnak. Ezek a molekulák szerkezetileg hasonlítanak a klorofillhoz, de abszorpciós spektrumuk a közeli infravörös tartományba tolódott el (700-1000 nm). Ez az adaptáció lehetővé teszi számukra, hogy olyan környezetekben is fotoszintetizáljanak, ahol a látható fény már elnyelődött, például mélyen a víz alatt vagy a talajban, ahol csak infravörös sugárzás áll rendelkezésre.
Ez a sokszínűség rávilágít arra, hogy míg a klorofill a a fotoszintézis központi eleme, az élet evolúciója során számos különböző pigmentrendszer alakult ki, amelyek optimalizálják a fényenergia hasznosítását a legkülönfélébb ökológiai fülkékben. A klorofill a alapvető szerepe mellett a kiegészítő pigmentek és a specifikus fénygyűjtő komplexek teszik lehetővé az élet számára, hogy a napenergia szinte minden elérhető formáját kiaknázza.
A klorofill a fluoreszcenciája és a fotoszintézis hatékonysága
Amikor a klorofill a molekula elnyel egy fotont és gerjesztett állapotba kerül, az elnyelt energia három fő úton távozhat belőle:
- Fotokémiai energiaátadás: Az energia továbbítódik a reakcióközpontba, és elindítja a fotoszintézist. Ez a kívánt és leghatékonyabb út.
- Hőleadás: Az energia hő formájában disszipálódik, különösen akkor, ha túl sok fény érkezik, és a fotoszintetikus apparátus telített.
- Fluoreszcencia: Az energia egy része alacsonyabb energiájú foton formájában újra kibocsátódik, általában vörös fényként.
A klorofill a fluoreszcenciája a gerjesztett állapotból az alapállapotba való visszatéréskor kibocsátott fény. Mivel a klorofill a főként vörös és kék fényt nyel el, a fluoreszcencia jellemzően a vörös spektrum tartományába esik, körülbelül 680-740 nm hullámhosszon. Ez a jelenség nem csupán érdekesség, hanem egy rendkívül fontos eszköz a fotoszintézis hatékonyságának és a növények stresszállapotának mérésére.
A fluoreszcencia mint indikátor
A klorofill a fluoreszcenciájának intenzitása és kinetikája közvetlen összefüggésben áll a fotoszintézis hatékonyságával. Ha a fotoszintetikus reakcióközpontok nyitottak (azaz készen állnak az elektronok fogadására), akkor az elnyelt fényenergia nagy része fotokémiai úton hasznosul, és a fluoreszcencia alacsony. Ha azonban a reakcióközpontok telítettek, vagy a növény stressz alatt áll (pl. vízhiány, hőstressz, túlzott fény), és a fotokémiai energiaátadás csökken, akkor a felesleges energia nagyobb része hővé alakul, és a fluoreszcencia is megnő. Ez a jelenség a Kautsky-effektus néven ismert.
A klorofill fluoreszcencia mérése lehetővé teszi a kutatók és agrárszakemberek számára, hogy:
- Mérjék a fotoszintézis kvantumhatékonyságát: Hány fotonból hány elektron transzportálódik.
- Értékeljék a növények stresszállapotát: A magasabb fluoreszcencia intenzitás gyakran stresszre utal.
- Optimalizálják a növénytermesztési körülményeket: Például a mesterséges megvilágítás intenzitását és spektrumát.
- Felmérjék az ökológiai rendszerek egészségét: Távoli érzékeléssel (pl. drónokról, műholdakról) monitorozható a vegetáció fotoszintetikus aktivitása.
A fluoreszcencia mérésének egyik legelterjedtebb módszere a pulzusamplitúdó modulált (PAM) fluorimetria. Ez a technika lehetővé teszi a fluoreszcencia gyors és roncsolásmentes mérését, és képes megkülönböztetni a különböző energiahasznosító útvonalakat (fotokémia, hőleadás, fluoreszcencia).
A fotoprotekció és a klorofill a
A klorofill a fluoreszcenciája mellett a növények rendelkeznek más mechanizmusokkal is a felesleges fényenergia kezelésére, azaz a fotoprotekcióra. Mint említettük, a karotinoidok kulcsszerepet játszanak ebben, elnyelve a túlzott energiát és hővé alakítva azt. Ez a non-fotokémiai kioltás (NPQ – Non-Photochemical Quenching) segít megakadályozni a fotoszintetikus apparátus károsodását, amelyet a reaktív oxigénfajták (ROS) okozhatnának a túlzott fényenergia hatására.
A klorofill a tehát nem csupán a fényenergia befogásáért felelős, hanem a fotoszintetikus rendszer „visszajelző mechanizmusaként” is funkcionál a fluoreszcencia révén. Ez a visszajelzés alapvető fontosságú a növények számára a változó fényviszonyokhoz való alkalmazkodásban és az optimális fotoszintetikus hatékonyság fenntartásában.
A klorofill a fluoreszcenciájának tanulmányozása mélyebb betekintést enged a fotoszintézis finomhangolt szabályozásába, és gyakorlati alkalmazásokat kínál a növények egészségének és produktivitásának monitorozásában.
A klorofill a és a modern kutatás: a jövő perspektívái

A klorofill a nem csupán egy jól ismert biológiai molekula, hanem a modern tudományos kutatás és technológiai fejlesztések egyik fókuszpontja is. A fotoszintézis alapvető mechanizmusainak mélyebb megértése, amelyben a klorofill a központi szerepet játszik, inspirálja a megújuló energiaforrások, a biotechnológia és a fenntartható mezőgazdaság innovációit.
Mesterséges fotoszintézis
Az egyik legizgalmasabb kutatási terület a mesterséges fotoszintézis. A tudósok arra törekednek, hogy utánozzák a természetes fotoszintézis hatékonyságát és mechanizmusait, különösen a víz bontását oxigénné és protonokká, valamint a szén-dioxid redukcióját üzemanyagokká (pl. hidrogén, metán, szén-monoxid) vagy szerves vegyületekké. A klorofill a fényelnyelő és elektronátadó tulajdonságai kulcsfontosságúak ezen a területen.
A kutatók szintetikus pigmenteket és katalizátorokat fejlesztenek, amelyek képesek a klorofill a funkcióját utánozni, elnyelve a napfényt és felhasználva azt kémiai reakciók hajtására. Ez a technológia óriási potenciállal rendelkezik a tiszta, megújuló energia előállításában, minimalizálva a fosszilis tüzelőanyagoktól való függőséget.
Biotechnológiai alkalmazások
A klorofill a és a fotoszintetikus apparátus ismerete számos biotechnológiai alkalmazáshoz vezet. Például:
- Alga bioreaktorok: Az algák, amelyek nagy mennyiségű klorofill a-t tartalmaznak, hatékonyan képesek szén-dioxidot megkötni és biomasszát termelni. Ez a biomassza felhasználható bioüzemanyagok (biodízel), élelmiszer-adalékanyagok, takarmányok vagy gyógyszerek előállítására.
- Növényi stressz monitorozás: A klorofill fluoreszcencia mérésére alapuló technológiák (ahogy korábban tárgyaltuk) lehetővé teszik a növények egészségi állapotának valós idejű monitorozását, segítve a termelőknek a termés optimalizálását és a stresszhatások megelőzését.
- Genetikai módosítás: A klorofill bioszintéziséért felelős gének módosításával a tudósok javíthatják a növények fotoszintetikus hatékonyságát, növelhetik a terméshozamot, vagy növelhetik a növények ellenálló képességét a környezeti stresszel szemben.
Környezeti és ökológiai kutatások
A klorofill a koncentrációjának mérése kulcsfontosságú indikátor az ökológiai rendszerek állapotának felmérésében. A vízi környezetben (óceánok, tavak) a klorofill a mennyisége közvetlenül arányos a fitoplankton biomasszájával, amely az elsődleges termelő a vízi táplálékláncokban. Műholdas távérzékeléssel a kutatók globális szinten monitorozhatják a fitoplankton virágzását, nyomon követhetik a klímaváltozás hatásait az óceánok produktivitására, és előrejelezhetik a halállományok mozgását.
A szárazföldi ökoszisztémákban a klorofill a tartalom a vegetáció vitalitására utal. A NDVI (Normalizált Differenciált Vegetációs Index), amely a vörös és a közeli infravörös fény visszaverődésének különbségén alapul, a klorofill a abszorpciós tulajdonságait használja fel a növényzet sűrűségének és egészségének becslésére. Ez a technika elengedhetetlen az erdőgazdálkodásban, a mezőgazdaságban és a globális szénciklus modellezésében.
A klorofill a nem csupán a fotoszintézis alappillére, hanem a modern tudomány inspirációja is, amely a fenntartható jövő megteremtéséhez vezető innovációk alapját képezi.
A klorofill a vizsgálata továbbra is a biológia, kémia és fizika határterületein zajlik, feltárva a molekula komplex kölcsönhatásait a környezetével és a fotoszintetikus apparátus más komponenseivel. Ez a folyamatos kutatás nemcsak elméleti tudásunkat bővíti, hanem gyakorlati megoldásokat is kínál a globális élelmezési és energiaellátási kihívásokra.
A klorofill a stabilitása és lebomlása a környezetben
A klorofill a, mint rendkívül fontos pigment, nemcsak a növényi sejtekben, hanem a környezetben is dinamikus változásokon megy keresztül a lebomlási folyamatok során. Ennek megértése kulcsfontosságú az ökológiai ciklusok, a tápanyag-újrahasznosítás és a környezeti szennyezés hatásainak tanulmányozásában.
A klorofill lebomlása a természetben
Amikor a növényi szövetek elpusztulnak (pl. lehullott levelek, elhalt algák), a klorofill a molekulák lebomlanak. Ez a folyamat többnyire enzimatikus úton történik, de fizikai és kémiai tényezők is befolyásolják, mint például a fény, a hőmérséklet, az oxigén és a pH. A lebomlás során a klorofill a különböző klorofill származékokká alakul át, amelyek közül néhányat a környezeti monitorozásban indikátorként használnak.
A lebomlási termékek közé tartoznak a feofitin és a feoforbid. A feofitin a klorofill a magnéziummentes formája, amely akkor keletkezik, amikor a magnéziumion kilép a porfirin gyűrűből savas körülmények között (pl. talajban, bomló biomasszában). A feoforbid a feofitin fitol farok nélküli változata. Ezek a pigmentek már nem képesek fotoszintézisre, és jelzik a fotoszintetikus anyag lebomlását.
A bomlási folyamatok során felszabaduló nitrogén és magnézium visszakerül a talajba vagy a vízi környezetbe, ahol más szervezetek (baktériumok, gombák) újrahasznosítják, ezzel zárva be a tápanyagciklusokat. Ez az újrahasznosítás alapvető fontosságú az ökoszisztémák termelékenységének fenntartásához.
Környezeti stressz és a klorofill a
A klorofill a mennyisége és stabilitása érzékeny indikátora a növények és algák környezeti stresszállapotának. A környezeti tényezők, mint például:
- Szennyező anyagok: Nehézfémek, peszticidek, levegőszennyezők.
- Vízhány: Aszály.
- Hőmérsékleti extrémumok: Fagy, hőség.
- Tápanyaghiány: Nitrogén, magnézium, vas hiánya.
- Túlzott fény: Fotoinhibíció.
mind befolyásolhatják a klorofill a szintézisét és lebomlását. Például a nitrogénhiány gátolja a klorofill szintézisét, ami a levelek sárgulásához (klorózis) vezet. A nehézfémek károsíthatják a klorofill molekulát vagy gátolhatják a fotoszintetikus enzimek működését, csökkentve a fotoszintetikus hatékonyságot.
A klorofill a változásainak mérése (pl. klorofill tartalom, klorofill a/b arány, fluoreszcencia) széles körben alkalmazott módszer a növények stresszreakcióinak tanulmányozására és a környezeti szennyezés hatásainak felmérésére. Ez a megközelítés lehetővé teszi a környezetvédelmi szakemberek számára, hogy korai előrejelzéseket tegyenek az ökoszisztémák állapotáról, és beavatkozásokat tervezzenek a káros hatások mérséklésére.
A klorofill a mint biomarker
A klorofill a nemcsak a földi és vízi ökoszisztémákban, hanem akár az űrkutatásban is releváns lehet. A távoli bolygók vagy exobolygók légkörének spektroszkópiai vizsgálata során a klorofill a abszorpciós spektrumára jellemző „vörös szél” (red edge) detektálása potenciálisan az élet jelenlétére utalhat. Ez a spektrális jelenség a fotoszintetikus organizmusok által visszavert közeli infravörös fény hirtelen növekedését jelenti a vörös fény elnyelése után.
Bár más pigmentek is léteznek, a klorofill a univerzális szerepe a földi fotoszintézisben azt sugallja, hogy hasonló pigmentek is kialakulhatnak más bolygókon, ha ott is a fényenergia a fő energiaforrás. Ez a „biosignature” keresése a asztrrobiológia egyik izgalmas területe.
A klorofill a tehát nem csupán egy molekula, hanem egy komplex biológiai rendszer központi eleme, amely folyamatosan kölcsönhatásban áll környezetével. Stabilitásának és lebomlásának dinamikája alapvető betekintést nyújt a földi élet ciklusába, az ökoszisztémák működésébe, és inspirációt ad a jövő tudományos és technológiai innovációihoz.







