
Az élet alapját képező molekula, a dezoxiribonukleinsav, vagy röviden DNS, az élőlények genetikai információjának hordozója. Ez a lenyűgöző makromolekula tartalmazza azokat az utasításokat, amelyek minden élőlény felépítéséhez, működéséhez és szaporodásához szükségesek. A DNS a sejtmagban található, és minden egyes sejtünkben, a baktériumoktól az emberig, gondosan elrendezve és védve őrzi az öröklődés titkait.
Minden élőlény egyedi jellemzői, mint például a szemszín, a magasság vagy éppen bizonyos betegségekre való hajlam, mind a DNS-ünkben kódolt információk eredményei. Ez a molekula tehát nem csupán egy kémiai vegyület, hanem az evolúció, az adaptáció és a biológiai sokféleség motorja is. Ahhoz, hogy megértsük az élet komplexitását, elengedhetetlen a DNS felépítésének és működésének alapos ismerete.
A DNS felfedezésének története
A DNS, mint az örökítőanyag, felfedezése nem egyetlen pillanat műve volt, hanem egy hosszú, kutatói generációkon átívelő tudományos utazás eredménye. A történet egészen a 19. század végéig nyúlik vissza, amikor a tudósok először kezdték megsejteni, hogy valamilyen molekula felelős lehet a tulajdonságok átadásáért.
Az első jelentős lépést Friedrich Miescher tette meg 1869-ben, amikor gennyes kötszerekből és lazacivarból izolált egy addig ismeretlen, foszfortartalmú, savas anyagot, amelyet „nukleinnek” nevezett el. Ez az anyag a sejtek magjában koncentrálódott, innen ered a neve is, és ez volt a DNS első azonosítása, bár Miescher ekkor még nem tudta annak valódi biológiai jelentőségét.
A 20. század elején Phoebus Levene orosz biokémikus tovább kutatta a nukleint, és felismerte, hogy az cukorból, foszfátcsoportból és nitrogéntartalmú bázisokból áll. Ő vezette be a nukleotid fogalmát, mint a DNS alapvető építőegységét, és azonosította a négy bázist: adenint (A), guanint (G), citozint (C) és timint (T). Levene azonban tévesen úgy vélte, hogy ezek a nukleotidok ismétlődő, egyszerű mintázatban követik egymást, ami kizárta volna, hogy a DNS hordozza a komplex genetikai információt.
Az 1940-es években Erwin Chargaff osztrák biokémikus jelentős áttörést ért el. Kísérletei során kimutatta, hogy a különböző fajok DNS-eiben a négy bázis aránya eltérő, de egy adott fajon belül mindig állandó. A legfontosabb felfedezése az volt, hogy a DNS-ben az adenin mennyisége mindig közel azonos a timin mennyiségével (A=T), és hasonlóképpen a guanin mennyisége is megegyezik a citozin mennyiségével (G=C). Ezeket az összefüggéseket ma Chargaff-szabályokként ismerjük, és kulcsfontosságúak voltak a DNS szerkezetének megfejtéséhez.
A valódi áttörés az 1950-es évek elején következett be, amikor Rosalind Franklin és Maurice Wilkins röntgendiffrakciós felvételeket készítettek a DNS-ről a londoni King’s College-ban. Franklin kristálytiszta képeket állított elő, amelyek egyértelműen utaltak a DNS spirális szerkezetére. Különösen az egyik, „Photo 51” néven ismert felvétel bizonyult döntőnek.
Ezekre az adatokra, valamint Chargaff szabályaira támaszkodva, James Watson és Francis Crick a Cambridge-i Egyetemen 1953-ban publikálták a DNS kettős spirál modelljét. Az ő modelljük elegánsan magyarázta, hogyan tárolódik az információ, és hogyan másolódik a DNS. A bázispárosodás (A-T és G-C) elve nemcsak a szerkezetet, hanem a genetikai információ átadásának mechanizmusát is megvilágította. Watson, Crick és Wilkins 1962-ben orvosi Nobel-díjat kapott ezért a korszakalkotó felfedezésért, Franklin sajnos addigra már elhunyt.
A DNS kettős spiráljának felfedezése nem csupán egy tudományos áttörés volt, hanem egy paradigmaváltás, amely örökre megváltoztatta az élet megértésének módját.
A DNS alapvető építőkövei: a nukleotidok
A DNS egy polimer, ami azt jelenti, hogy számos kisebb egységből, az úgynevezett monomerekből épül fel. Ezek a monomerek a nukleotidok. Minden egyes nukleotid három alapvető részből áll, amelyek kémiai kötésekkel kapcsolódnak egymáshoz, és együttesen alkotják a DNS komplex szerkezetét.
Az első komponens egy foszfátcsoport. Ez egy foszforatomot tartalmaz, amelyet négy oxigénatom vesz körül. A foszfátcsoport felelős a DNS molekula negatív töltéséért, és kulcsszerepet játszik a nukleotidok összekapcsolódásában, létrehozva a DNS gerincét.
A második rész egy öt szénatomos cukor, a dezoxiribóz. Nevét onnan kapta, hogy a ribózhoz képest hiányzik róla egy oxigénatom (pontosabban a 2′ szénatomon hidrogénatom található hidroxilcsoport helyett). Ez a kis különbség rendkívül fontos a DNS stabilitása szempontjából, mivel a dezoxiribóz stabilabb, mint a ribóz, ami az RNS-ben található.
A harmadik és talán legváltozatosabb komponens a nitrogéntartalmú bázis. Ezek a heterociklusos vegyületek tartalmazzák a genetikai információt. Négyféle nitrogéntartalmú bázis létezik a DNS-ben: adenin (A), guanin (G), citozin (C) és timin (T). Ezek két fő csoportba sorolhatók kémiai szerkezetük alapján:
- Purinok: Két gyűrűből álló szerkezetek. Ide tartozik az adenin és a guanin.
- Pirimidinek: Egy gyűrűből álló szerkezetek. Ide tartozik a citozin és a timin.
A nukleotidok tehát úgy épülnek fel, hogy a dezoxiribóz cukor 1′ szénatomjához kapcsolódik a nitrogéntartalmú bázis, míg az 5′ szénatomjához a foszfátcsoport. Amikor a nukleotidok egymáshoz kapcsolódnak, a foszfátcsoport egy másik nukleotid 3′ szénatomjához kötődik, létrehozva egy foszfodiészter kötést. Ez az ismétlődő kötés képezi a DNS-szál gerincét, ahol a foszfát- és cukormolekulák váltakozva követik egymást.
Ez a „foszfát-cukor gerinc” rendkívül stabil, és a nitrogéntartalmú bázisok a gerinc belső felére, a kettős spirál középpontja felé mutatnak. Ez az elrendezés biztosítja, hogy a genetikai információ, amelyet a bázisok sorrendje kódol, védve legyen a környezeti hatásoktól, miközben mégis hozzáférhető marad a sejt számára a replikáció és transzkripció során.
A DNS kettős spiráljának szerkezete
A DNS legismertebb és legikonikusabb formája a kettős spirál, amelyet Watson és Crick írt le először. Ez a szerkezet nem csupán esztétikailag lenyűgöző, hanem funkcionálisan is tökéletesen alkalmas a genetikai információ tárolására és átadására. Két hosszú polinukleotid-láncból áll, amelyek egymás köré tekeredve egy spirális alakzatot alkotnak.
Minden egyes DNS-lánc egy cukor-foszfát gerincből áll, amely a nukleotidok dezoxiribóz cukor- és foszfátcsoportjaiból épül fel, foszfodiészter kötésekkel összekapcsolva. Ezek a gerincek a kettős spirál külső részét alkotják, mintegy védőburkot képezve. A nitrogéntartalmú bázisok (adenin, guanin, citozin, timin) pedig a gerinc belső oldalára néznek, a spirál középpontja felé.
A két DNS-szálat a bázisok közötti specifikus hidrogénkötések tartják össze. Ez a bázispárosodás rendkívül specifikus és kulcsfontosságú a DNS funkciójában. Az adenin (A) mindig a timinnel (T) párosodik, két hidrogénkötéssel, míg a guanin (G) mindig a citozinnal (C) párosodik, három hidrogénkötéssel. Ezt az elvet nevezzük komplementer bázispárosodásnak. Ennek köszönhetően a két szál egymás tükörképe, vagyis ha ismerjük az egyik szál bázissorrendjét, azonnal tudjuk a másikét is.
A két szál nem párhuzamosan fut, hanem antiparallel módon. Ez azt jelenti, hogy az egyik szál az 5′ végétől a 3′ végéig halad, míg a másik szál ellentétes irányban, a 3′ végétől az 5′ végéig. Az „5′ vég” a dezoxiribóz cukor ötödik szénatomján lévő foszfátcsoportra utal, míg a „3′ vég” a harmadik szénatomon lévő hidroxilcsoportra. Ez az antiparallel elrendezés alapvető fontosságú a DNS replikációja és transzkripciója során.
A kettős spirál nem egy sima henger, hanem jellegzetes hornyokat (barázdákat) mutat. Kétféle horony létezik: a nagy horony (major groove) és a kis horony (minor groove). Ezek a hornyok fontosak, mert lehetővé teszik a fehérjék számára, hogy felismerjék és specifikusan kötődjenek a DNS-hez anélkül, hogy fel kellene tekerniük a spirált. Ez a felismerés kritikus a génexpresszió szabályozásában.
A DNS stabilitása több tényezőnek köszönhető. A foszfát-cukor gerinc kovalens kötései erősek. A bázisok közötti hidrogénkötések önmagukban viszonylag gyengék, de a több millió bázispár együttesen hatalmas stabilitást biztosít. Emellett a bázisok egymásra torlódása (stacking interaction) a hidrofób kölcsönhatások révén szintén hozzájárul a szerkezet stabilitásához. Ez a robusztus felépítés teszi lehetővé, hogy a DNS biztonságosan tárolja az élet kódját.
| Jellemző | Leírás |
|---|---|
| Két polinukleotid-lánc | Egyszerre két szál alkotja a spirált. |
| Cukor-foszfát gerinc | A láncok külső részét alkotó, stabil váz. |
| Nitrogéntartalmú bázisok | A spirál belseje felé mutatnak, információt hordoznak. |
| Komplementer bázispárosodás | A-T (2 H-kötés), G-C (3 H-kötés). |
| Antiparallel szálak | A két szál ellentétes irányban fut (5’→3′ és 3’→5′). |
| Nagy és kis horony | A spirál felületén található barázdák a fehérje-kötéshez. |
| Stabilitás | Kovalens kötések, hidrogénkötések és bázis stacking révén. |
A DNS becsomagolása: a spiráltól a kromoszómáig
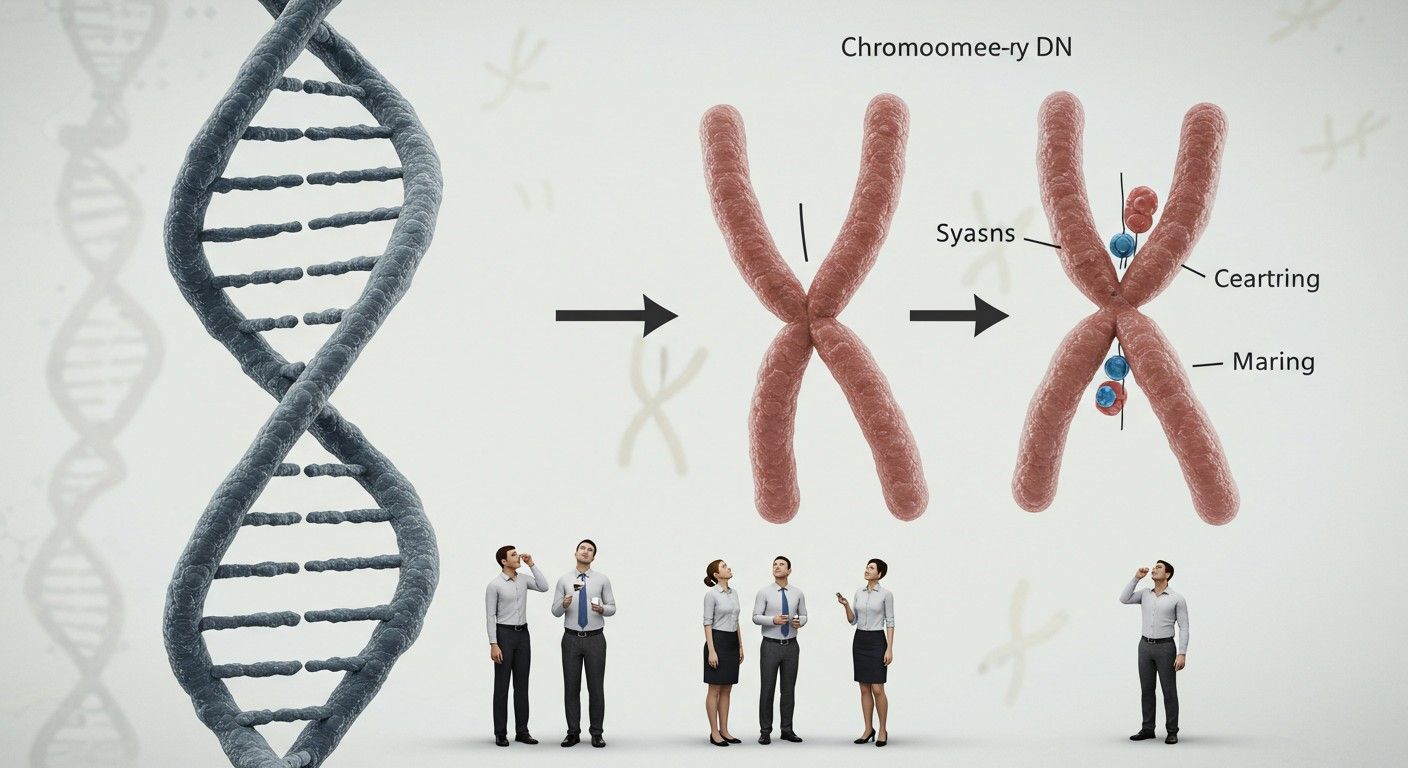
Az emberi sejtmagban található DNS hossza elképesztő. Egyetlen emberi sejt DNS-e kinyújtva körülbelül 2 méter hosszú lenne. Ahhoz, hogy ez a hatalmas molekula elférjen a mindössze néhány mikrométer átmérőjű sejtmagban, rendkívül hatékony és precíz becsomagolásra van szüksége. Ez a folyamat nem csupán a helytakarékosságot szolgálja, hanem a génexpresszió szabályozásában is kulcsfontosságú szerepet játszik.
Az első szintű csomagolásban a DNS a hisztonok nevű fehérjék köré tekeredik. A hisztonok kis méretű, pozitív töltésű fehérjék, amelyek erősen kötődnek a DNS negatív töltésű foszfátcsoportjaihoz. Öt fő típusú hiszton létezik (H1, H2A, H2B, H3, H4), amelyek közül négy (H2A, H2B, H3, H4) két-két példánya egy nyolc fehérjéből álló hiszton oktamert alkot.
A DNS körülbelül 147 bázispárja tekeredik rá egy hiszton oktamerre, mintegy 1,65-ször körbevéve azt. Ez a szerkezet a nukleoszóma, amely a kromatin alapvető ismétlődő egysége, és „gyöngyök a fonalon” megjelenést kölcsönöz a DNS-nek mikroszkóp alatt. A H1 hiszton a nukleoszóma és a következő nukleoszóma közötti linker DNS-hez kötődik, stabilizálva a szerkezetet.
A nukleoszómák tovább tömörödnek, létrehozva a kromatin nevű komplexet. A kromatin két fő formában létezik:
- Eukromatin: Lazább szerkezetű, genetikailag aktív régiókat tartalmaz. Ez a DNS hozzáférhető a transzkripcióhoz, vagyis a gének kifejeződéséhez.
- Heterokromatin: Sűrűbben csomagolt, genetikailag inaktív régiókat tartalmaz. Ez a DNS általában nem transzkribálódik, és gyakran ismétlődő szekvenciákat, például centroméreket és teloméreket foglal magában.
A kromatin tovább spirálizálódik és kondenzálódik, különösen a sejtosztódás, azaz a mitózis során. Ekkor válnak láthatóvá a kromoszómák, amelyek a DNS legmagasabb szintű csomagolási formái. Minden kromoszóma egyetlen hosszú DNS-molekulát tartalmaz, amely szorosan feltekeredett és kondenzálódott. Az emberi test sejtjeiben 23 pár, azaz 46 kromoszóma található.
A kromoszómák szerkezetében két fontos régió emelhető ki: a centromér és a telomerek. A centromér a kromoszóma középső, elvékonyodott része, amely a sejtosztódás során a testvérkromatidák összekapcsolásáért és a mikrotubulusokhoz való rögzüléséért felelős. A telomerek a kromoszómák végein található ismétlődő DNS-szekvenciák, amelyek védelmet nyújtanak a DNS-nek a lebomlástól és a fúziótól, valamint szerepet játszanak a sejtöregedésben.
Ez a komplex csomagolási mechanizmus biztosítja, hogy a hatalmas mennyiségű genetikai információ ne csak elférjen a sejtmagban, hanem rendezetten és szabályozottan hozzáférhető is legyen, amikor arra szükség van. A kromatin szerkezetének módosítása, például a hisztonok kémiai módosításai (acetiláció, metiláció), az epigenetika területén kulcsfontosságúak, mivel befolyásolják a gének kifejeződését anélkül, hogy megváltoztatnák a DNS szekvenciáját.
A DNS csomagolása nem csupán fizikai kényszer, hanem egy dinamikus folyamat, amely finomhangolja a génexpressziót és alapvető szerepet játszik a sejtek működésében és fejlődésében.
A molekuláris biológia központi dogmája
A molekuláris biológia központi dogmája egy alapvető koncepció, amelyet Francis Crick fogalmazott meg 1957-ben, majd 1970-ben részletesebben is kifejtett. Ez a dogma leírja a genetikai információ áramlásának fő irányát a biológiai rendszerekben: DNS-ből RNS-be, RNS-ből pedig fehérjébe. Ez az elv a mai napig az egyik legfontosabb alapja a modern biológiai kutatásoknak, és magyarázatot ad arra, hogyan valósul meg a genetikai információ a sejtekben.
A központi dogma három fő folyamatot foglal magában:
- Replikáció (DNS → DNS): A DNS képes önmagát másolni, ami elengedhetetlen a sejtek osztódásához és az örökítőanyag generációról generációra történő pontos átadásához. Ez a folyamat biztosítja, hogy minden utódsejt megkapja a teljes genetikai információt.
- Transzkripció (DNS → RNS): A genetikai információt a DNS-ről átírják egy rokon molekulára, az RNS-re (ribonukleinsavra). Ez a lépés egyfajta „munka másolatot” hoz létre a DNS egy adott szakaszáról, egy génről, amelyre a sejtnek éppen szüksége van.
- Transzláció (RNS → Fehérje): Az RNS-ben tárolt információt lefordítják a fehérjék aminosavsorrendjére. A fehérjék a sejtek „munkásai”, amelyek szinte minden életfolyamatban részt vesznek, legyen szó szerkezeti elemekről, enzimekről vagy jelátviteli molekulákról.
Ez a három folyamat egymásra épül, és szigorúan szabályozott, biztosítva a génexpresszió precíz irányítását. Bár a dogma alapvetően egyirányúnak írja le az információáramlást, később felfedeztek néhány kivételt, például a reverz transzkripciót, ahol az RNS információja visszaíródik DNS-be (például retrovírusok, mint a HIV esetében). Azonban az alapvető elv, miszerint a DNS a genetikai információ végső tárháza, és a fehérjék a funkcionális termékek, változatlan maradt.
A replikáció során a DNS kettős spirálja széttekeredik, és mindkét szál mintaként szolgál egy új, komplementer szál szintéziséhez. Ennek eredményeként két azonos DNS-molekula jön létre, mindegyik egy eredeti és egy újonnan szintetizált szállal (ez a félig konzervatív replikáció elve).
A transzkripció során egy specifikus enzim, az RNS-polimeráz, a DNS egy génjét RNS-sé írja át. Az így keletkező RNS-t hírvivő RNS-nek (mRNS) nevezzük, ha fehérjét kódol, de léteznek más típusú RNS-ek is, mint például a transzfer RNS (tRNS) és a riboszomális RNS (rRNS), amelyek a transzlációban játszanak szerepet.
Végül, a transzláció a riboszómákon megy végbe. Az mRNS-ben lévő információt, amely kodonok formájában (három nukleotidból álló egységek) van kódolva, a tRNS molekulák „fordítják le” aminosavakra. Minden tRNS egy specifikus aminosavat szállít, és egy antikodonnal rendelkezik, amely komplementer az mRNS kodonjával. Az aminosavak összekapcsolódnak, így alakul ki a fehérje polipeptid lánca.
Ez a precíz és összehangolt rendszer biztosítja, hogy a genetikai információ pontosan kifejeződjön, és lehetővé teszi a sejtek számára, hogy a környezeti változásokra reagálva szabályozzák génjeik működését. A központi dogma megértése alapvető fontosságú a genetikai betegségek, a gyógyszerfejlesztés és a biotechnológia számos területén.
A DNS replikációja: az örökítőanyag másolása
A DNS replikációja az a biológiai folyamat, amely során a DNS molekula önmagát másolja, két azonos DNS-molekulát hozva létre. Ez a mechanizmus létfontosságú az élet fenntartásához, mivel biztosítja, hogy minden sejtosztódás során az utódsejtek pontosan ugyanazt a genetikai információt kapják meg, mint az anyasejt. Ez a folyamat rendkívül precíz és szabályozott, minimalizálva a hibák lehetőségét.
A replikáció félig konzervatív módon történik, ahogyan azt Matthew Meselson és Franklin Stahl igazolta 1958-ban. Ez azt jelenti, hogy minden újonnan szintetizált DNS-molekula egy eredeti (templát) és egy újonnan szintetizált szálból áll. Az eredeti kettős spirál két szálra válik szét, és mindegyik szál mintaként szolgál egy új komplementer szál szintéziséhez.
A replikáció egy sor speciális enzim összehangolt munkáját igényli:
- Helikáz: Ez az enzim felelős a DNS kettős spiráljának feltekeréséért és a hidrogénkötések felbontásáért a két szál között, létrehozva egy replikációs villát.
- Topoisomeráz: Mivel a helikáz a DNS-t feltekeri, a spirál előtt feszültség keletkezik. A topoisomerázok enyhítik ezt a feszültséget, megakadályozva a DNS túlzott tekercselődését és károsodását.
- SSBP (Single-Strand Binding Proteins): Ezek a fehérjék a szétválasztott DNS-szálakhoz kötődnek, megakadályozva azok visszatekeredését, amíg az új szál szintézise folyik.
- Primáz: A DNS-polimeráz nem képes teljesen új DNS-szálat szintetizálni a semmiből. Ezért a primáz egy rövid RNS-szakasz, az úgynevezett primer szintézisével indítja el a folyamatot, amelyhez a DNS-polimeráz hozzákapcsolódhat.
- DNS-polimeráz III: Ez a fő enzim felelős az új DNS-szálak szintéziséért. Az eredeti templát szál mentén haladva, komplementer nukleotidokat illeszt be, és foszfodiészter kötésekkel kapcsolja össze őket. A DNS-polimeráz III csak az 5′ → 3′ irányban képes szintetizálni.
- DNS-polimeráz I: Ez az enzim eltávolítja az RNS primereket, és helyükre DNS-nukleotidokat szintetizál.
- Ligáz: A ligáz enzim összekapcsolja az újonnan szintetizált DNS-szakaszokat, létrehozva a folytonos DNS-szálat.
Mivel a DNS-polimeráz csak az 5′ → 3′ irányban képes dolgozni, és a két templát szál antiparallel, a replikáció két különböző módon zajlik a replikációs villán belül:
- Vezető szál (leading strand): Ezen a szálon a szintézis folyamatosan, a replikációs villa mozgásának irányában történik, egyetlen primer felhasználásával.
- Késlekedő szál (lagging strand): Ezen a szálon a szintézis szakaszosan, a replikációs villa mozgásával ellentétes irányban történik. Ennek eredményeként rövid DNS-szakaszok, az úgynevezett Okazaki-fragmentumok jönnek létre, amelyek mindegyike egy RNS primerrel indul. Ezeket az Okazaki-fragmentumokat később a DNS-polimeráz I és a ligáz kapcsolja össze.
A replikáció pontossága rendkívül magas, köszönhetően a DNS-polimerázok beépített hibajavító (proofreading) képességének. Ha egy hibás nukleotid illeszkedik be, az enzim azonnal felismeri, eltávolítja, és helyére a megfelelő bázist építi be. Ez a mechanizmus drámaian csökkenti a mutációk arányát, biztosítva a genetikai információ integritását.
A DNS replikációja egy csodálatosan összetett és precíz gépezet, amely garantálja az élet folytonosságát és a genetikai örökség hűséges átadását minden sejtosztódás során.
Transzkripció: a genetikai üzenet átírása RNS-re
A transzkripció az a folyamat, amely során a genetikai információ a DNS-ről RNS-re íródik át. Ez a lépés a génexpresszió első és egyik legfontosabb szabályozási pontja, amely lehetővé teszi a sejt számára, hogy csak azokra a génekre „kapcsoljon be”, amelyekre éppen szüksége van. A transzkripció során nem az egész DNS másolódik le, hanem csak a gének specifikus szakaszai.
A transzkripciót egy enzim, az RNS-polimeráz végzi. Ez az enzim képes felismerni a DNS-en található speciális szekvenciákat, az úgynevezett promótereket, amelyek jelzik a gén kezdetét. A promóterekhez kötődve az RNS-polimeráz feltekercseli a DNS kettős spirálját, és szétválasztja a két szálat, létrehozva egy rövid replikációs buborékot.
Ezt követően az RNS-polimeráz a DNS egyik szálát (a templát szálat) mintaként használja, és komplementer RNS-nukleotidokat épít be az 5′ → 3′ irányban. Fontos különbség a DNS-hez képest, hogy az RNS-ben a timin (T) helyett uracil (U) található. Tehát, ha a DNS templát szálán adenin (A) van, az RNS-ben uracil (U) épül be; ha guanin (G) van, citozin (C) épül be, és fordítva. A citozin (C) helyére guanin (G) jön, a timin (T) helyére pedig adenin (A).
A transzkripció addig folytatódik, amíg az RNS-polimeráz el nem ér egy másik speciális DNS-szekvenciát, a terminátort. Ez jelzi a gén végét, és a polimeráz ekkor leválik a DNS-ről, felszabadítva az újonnan szintetizált RNS-molekulát.
Három fő típusú RNS molekula képződik a transzkripció során, amelyek mindegyike alapvető szerepet játszik a génexpresszióban:
- Hírvivő RNS (mRNS): Ez hordozza a fehérjék szintéziséhez szükséges genetikai kódot a DNS-től a riboszómákig.
- Transzfer RNS (tRNS): Ezek a molekulák felelősek az aminosavak szállításáért a riboszómákhoz a fehérjeszintézis során.
- Riboszomális RNS (rRNS): Ezek a molekulák alkotják a riboszómák szerkezeti és katalitikus magját, ahol a fehérjeszintézis zajlik.
Az eukarióta sejtekben a transzkripció után az újonnan szintetizált RNS, az úgynevezett pre-mRNS, még további módosításokon esik át, mielőtt elhagyhatná a sejtmagot és részt vehetne a fehérjeszintézisben. Ezek a poszt-transzkripciós módosítások a következők:
- Sapkázás (capping): Az mRNS 5′ végére egy speciális guanozin nukleotid, a 7-metilguanozin sapka kerül. Ez védi az mRNS-t a lebomlástól, és segíti a riboszómához való kötődését.
- Poliadeniláció (polyadenylation): Az mRNS 3′ végére egy hosszú adenin nukleotid sorozat, az úgynevezett poli-A farok kerül. Ez szintén növeli az mRNS stabilitását és szerepet játszik a sejtmagból való exportálásában.
- Splicing (splicing): Az eukarióta gének gyakran tartalmaznak nem kódoló szakaszokat, az úgynevezett intronokat, amelyek megszakítják a kódoló régiókat, az exonokat. A splicing során az intronokat kivágják, és az exonokat pontosan összekapcsolják, létrehozva az érett mRNS-t. Az alternatív splicing révén egyetlen génből többféle fehérje is létrejöhet.
Ezek a módosítások biztosítják, hogy az mRNS stabil, funkcionális és megfelelően fordítható legyen fehérjévé. A transzkripció szabályozása rendkívül összetett, és számos transzkripciós faktor, erősítő és gátló szekvencia vesz részt benne, lehetővé téve a sejtek számára a génexpresszió finomhangolását a különböző fejlődési stádiumokban és környezeti körülmények között.
Transzláció: az RNS-ből fehérje lesz

A transzláció, vagy más néven fehérjeszintézis, az a kritikus biológiai folyamat, amely során a genetikai információ, amelyet a hírvivő RNS (mRNS) hordoz, lefordítódik egy specifikus aminosavsorrendű fehérjévé. Ez a lépés a molekuláris biológia központi dogmájának utolsó állomása, és a sejt működéséhez szükséges összes fehérje előállításáért felelős.
A transzláció fő helyszíne a riboszóma, egy komplex molekuláris gép, amely riboszomális RNS-ből (rRNS) és fehérjékből áll. A riboszómák két alegységből, egy nagy és egy kis alegységből állnak, amelyek csak a fehérjeszintézis megkezdésekor kapcsolódnak össze.
A genetikai információ az mRNS-ben kodonok formájában van kódolva. Egy kodon három egymás utáni nukleotidból áll, és minden kodon egy specifikus aminosavat határoz meg, vagy egy stop jelet ad. A genetikai kód degenerált, ami azt jelenti, hogy több különböző kodon is kódolhatja ugyanazt az aminosavat, de minden kodon csak egyetlen aminosavat specifikál.
A transzláció folyamata három fő szakaszra bontható:
- Iniciáció (kezdés): A folyamat az mRNS 5′ végén található start kodonnal (AUG) kezdődik, amely a metionin aminosavat kódolja (eukariótákban formilmetionin prokariótákban). A kis riboszóma alegység, a start kodonhoz kötődő iniciátor tRNS (amely a metionint szállítja) és az mRNS összekapcsolódik, majd ehhez csatlakozik a nagy riboszóma alegység.
- Elongáció (lánchosszabbítás): Ebben a szakaszban a riboszóma az mRNS-en halad, kodonról kodonra. Minden egyes kodonhoz egy komplementer antikodonnal rendelkező transzfer RNS (tRNS) molekula kötődik. A tRNS-ek speciális aminosavakat szállítanak. Amikor egy tRNS bekerül a riboszóma „A” (aminosav) helyére, az aminosav összekapcsolódik a növekvő polipeptid lánccal egy peptidkötés révén. Ezután a riboszóma elmozdul egy kodonnal, a tRNS áthelyeződik a „P” (peptid) helyre, majd a „E” (kilépés) helyen távozik.
- Termináció (befejezés): A folyamat akkor ér véget, amikor a riboszóma eléri az mRNS-en található valamelyik stop kodont (UAA, UAG, UGA). Nincs olyan tRNS, amely ezekhez a kodonokhoz kötődne. Ehelyett felszabadító faktorok (release factors) kötődnek a stop kodonhoz, ami a polipeptid lánc leválását eredményezi a tRNS-ről és a riboszóma alegységeinek szétválását.
Az újonnan szintetizált polipeptid lánc ezután elkezdi felvenni specifikus háromdimenziós szerkezetét, azaz feltekeredik. Ez a feltekeredés spontán módon történhet, de gyakran segítő fehérjék, az úgynevezett chaperonok is részt vesznek benne. A fehérje helyes térbeli szerkezete elengedhetetlen a funkciójához. Néhány fehérje azonnal funkcionális, míg mások további módosításokon esnek át (pl. glikoziláció, hasítás) a funkciójuk eléréséhez.
A transzláció rendkívül hatékony folyamat. Egy mRNS-molekulán egyszerre több riboszóma is dolgozhat, egy poliszómát vagy poliriboszómát alkotva, ami lehetővé teszi számos fehérje gyors előállítását ugyanabból a genetikai üzenetből. Ez a komplex gépezet biztosítja, hogy a sejt képes legyen gyorsan és pontosan reagálni a változó igényekre, és előállítsa az összes szükséges funkcionális molekulát az élet fenntartásához.
A genetikai kód: univerzalitás és degeneráció
A genetikai kód az a szabályrendszer, amely meghatározza, hogyan fordítódnak le a DNS-ben és RNS-ben tárolt nukleotid-szekvenciák a fehérjék aminosavsorrendjére. Ez az alapvető kód az élet minden ismert formájában szinte azonos, ami az evolúció egyik legmeggyőzőbb bizonyítéka.
A genetikai kód alapja a triplet kód. Ez azt jelenti, hogy minden egyes aminosavat három egymást követő nukleotidból álló egység, az úgynevezett kodon határoz meg. Mivel négyféle nukleotid (A, U, G, C az RNS-ben) létezik, 4³ = 64 lehetséges kodon kombináció létezik. Mivel azonban csak 20 féle standard aminosav van, ez azt jelenti, hogy a kód degenerált (vagy redundáns).
A degeneráció azt jelenti, hogy a legtöbb aminosavat több különböző kodon is kódolja. Például a leucin aminosavat hat különböző kodon is specifikálja (UUA, UUG, CUU, CUC, CUA, CUG). Ez a redundancia előnyös lehet, mivel csökkenti a pontmutációk káros hatásait. Ha egyetlen nukleotid megváltozik egy kodonban, a mutáció attól még kódolhatja ugyanazt az aminosavat, vagy egy hasonló tulajdonságú aminosavat, minimalizálva a fehérje funkciójának romlását.
A 64 kodon közül 61 aminosavakat kódol, míg a maradék három kodon, az UAA, UAG és UGA, úgynevezett stop kodonok. Ezek nem kódolnak aminosavat, hanem a fehérjeszintézis befejezését jelzik. A start kodon, amely a fehérjeszintézis kezdetét jelöli, az AUG, amely a metionin aminosavat is kódolja.
A genetikai kód egyik legmegdöbbentőbb tulajdonsága az univerzalitása. Szinte minden élőlényben, a baktériumoktól az emberig, ugyanazok a kodonok ugyanazokat az aminosavakat kódolják. Ez azt jelenti, hogy egy bakteriális gén, amelyet egy emberi sejtbe ültetnek, ugyanazt a fehérjét fogja termelni, és fordítva. Ez az univerzalitás alapvető bizonyíték az élet közös eredetére, és lehetővé teszi a biotechnológiai alkalmazásokat, mint például az inzulin termelése baktériumokban.
Bár a kód nagyrészt univerzális, léteznek apró kivételek, különösen a mitokondriális DNS-ben és néhány mikroorganizmusban. Például az emberi mitokondriumokban az UGA kodon triptofánt kódol stop jel helyett, és az AUA kodon metionint kódol izoleucin helyett. Ezek a kisebb eltérések azonban nem vonják kétségbe a kód alapvető univerzalitását.
A genetikai kód jellemzői összefoglalva:
- Triplet kód: Három nukleotid alkot egy kodont.
- Degenerált/Redundáns: Egy aminosavat több kodon is kódolhat.
- Univerzális: Szinte minden élőlényben azonos.
- Nem átfedő: A kodonok egymás után, átfedés nélkül olvashatók.
- Vesszőmentes: Nincsenek üres helyek vagy elválasztó jelek a kodonok között.
- Start és Stop kodonok: Jelzik a fehérjeszintézis kezdetét és végét.
Ez a kódolási rendszer rendkívül hatékony és robusztus, biztosítva, hogy a genetikai információ pontosan és megbízhatóan átíródjon funkcionális fehérjékké, amelyek az élet minden aspektusát irányítják.
Gépek: az öröklődés egységei
A gének az öröklődés alapvető fizikai és funkcionális egységei. Ezek a DNS specifikus szakaszai, amelyek tartalmazzák azokat az információkat, amelyek szükségesek egy RNS-molekula (és általában egy fehérje) szintéziséhez. A gének határozzák meg az élőlények tulajdonságait és jellemzőit, a hajszíntől kezdve a betegségekre való hajlamig.
Egy gén nem csupán a fehérjét kódoló szekvenciából áll. Sokkal komplexebb felépítése van, amely magában foglalja a kódoló régiókat és a nem kódoló, szabályozó régiókat is. Az eukarióta génekben a kódoló régiók gyakran megszakadnak nem kódoló szakaszokkal:
- Exonok: Ezek a gének kódoló részei, amelyek az érett mRNS-ben maradnak, és végül fehérjékké fordítódnak.
- Intronok: Ezek a gének nem kódoló részei, amelyek a transzkripció után kivágásra kerülnek (splicing révén), és nem jelennek meg az érett mRNS-ben. Bár nem kódolnak fehérjét, fontos szabályozó funkcióik lehetnek, vagy hozzájárulhatnak az alternatív splicinghez.
A gének működését számos szabályozó szekvencia befolyásolja, amelyek nem feltétlenül a gén kódoló részén belül helyezkednek el, de hatással vannak annak expressziójára:
- Promóterek: A gén elején található DNS-szekvenciák, amelyekhez az RNS-polimeráz és más transzkripciós faktorok kötődnek, elindítva a transzkripciót.
- Enhancerek (erősítők): Olyan távolabbi DNS-szekvenciák, amelyekhez specifikus aktivátor fehérjék kötődnek, és növelik a gén transzkripciójának sebességét.
- Szilencerek (gátlók): Az enhancerekhez hasonlóan távoli szekvenciák, de ezekhez represszor fehérjék kötődnek, amelyek csökkentik vagy gátolják a gén transzkripcióját.
A génexpresszió szabályozása rendkívül összetett és dinamikus folyamat. A sejtek képesek „be- és kikapcsolni” a géneket a fejlődési stádium, a környezeti ingerek vagy a sejt típusának megfelelően. Ez a szabályozás történhet a transzkripció szintjén (pl. a promóterek aktivitásának módosításával), a poszt-transzkripciós szinten (pl. az mRNS stabilitásának befolyásolásával), a transzlációs szinten (pl. a fehérjeszintézis sebességének szabályozásával) vagy a poszt-transzlációs szinten (pl. a fehérjék módosításával).
Az emberi genomban körülbelül 20 000-25 000 fehérjét kódoló gén található, de a teljes DNS-nek csak kis része (kb. 1-2%) kódol fehérjét. A fennmaradó, úgynevezett nem kódoló DNS, korábban „hulladék DNS-nek” tartották, ma már tudjuk, hogy számos fontos szabályozó funkciót lát el, például mikroRNS-eket, hosszú nem kódoló RNS-eket kódol, és szerepet játszik a kromatin szerkezetének fenntartásában.
A gén fogalma folyamatosan fejlődik a tudomány előrehaladtával. Ma már tudjuk, hogy a gének nem feltétlenül diszkrét, elkülönült egységek, hanem funkcionális régiók, amelyek átfedhetik egymást, vagy komplex kölcsönhatásban állhatnak más szabályozó elemekkel. A gének és a génexpresszió megértése alapvető fontosságú a biológia, az orvostudomány és a biotechnológia számára, lehetővé téve a betegségek mechanizmusainak megértését és új terápiák kifejlesztését.
A gének nem csupán a genetikai örökség passzív hordozói, hanem dinamikus egységek, amelyek működésükkel és szabályozásukkal formálják az életet és annak sokféleségét.
Mutációk: változások a DNS szekvenciájában
A mutációk a DNS nukleotid szekvenciájában bekövetkező tartós változások. Ezek a változások lehetnek olyan aprók, mint egyetlen bázispár megváltozása, vagy olyan nagyméretűek, mint egy egész kromoszóma darabjának átrendeződése. Bár a DNS replikációja rendkívül pontos, és a sejt számos hibajavító mechanizmussal rendelkezik, a mutációk elkerülhetetlenül bekövetkeznek. Ezek a változások az evolúció alapvető mozgatórugói, de számos betegség, köztük a rák, hátterében is állhatnak.
A mutációk okai lehetnek spontánok vagy indukáltak:
- Spontán mutációk: Ezek a sejt normális metabolikus folyamatai során keletkeznek, például a DNS replikációja során elkövetett hibák, vagy a bázisok kémiai instabilitása miatt. Ilyen lehet a tautomerizáció, ahol a bázisok ritka izomer formái téves párosodást okozhatnak.
- Indukált mutációk: Ezeket külső tényezők, úgynevezett mutagének okozzák. A mutagének lehetnek fizikai (pl. UV sugárzás, röntgensugárzás), kémiai (pl. etil-metánszulfonát, interkaláló szerek) vagy biológiai (pl. vírusok, transzpozonok) ágensek.
A mutációk típusai a DNS szintjén:
- Pontmutációk: Egyetlen nukleotid megváltozása.
- Szubsztitúció (csere): Egy bázispár cseréje egy másikra.
- Tranzíció: Purin purinra (A→G, G→A) vagy pirimidin pirimidinre (C→T, T→C) cserélődik.
- Transzverzió: Purin pirimidinre vagy pirimidin purinra cserélődik (A→C, A→T stb.).
- Inszerció (beillesztés): Egy vagy több nukleotid beillesztése a szekvenciába.
- Deléció (törlés): Egy vagy több nukleotid eltávolítása a szekvenciából.
- Szubsztitúció (csere): Egy bázispár cseréje egy másikra.
- Kromoszóma-mutációk: Nagyobb léptékű változások a kromoszómák szerkezetében vagy számában.
- Duplikáció: Egy kromoszóma-szakasz megkettőződése.
- Inverzió: Egy kromoszóma-szakasz megfordulása.
- Transzlokáció: Egy kromoszóma-szakasz áthelyeződése egy másik kromoszómára.
- Aneuploidia: A kromoszómák számának megváltozása (pl. Down-szindróma, ahol egy extra 21-es kromoszóma van).
A mutációk következményei a fehérje szintjén változatosak lehetnek:
- Csendes mutáció (silent mutation): A kodon megváltozik, de a degenerált genetikai kód miatt még mindig ugyanazt az aminosavat kódolja. Nincs hatása a fehérjére.
- Hibás értelmű mutáció (missense mutation): A kodon megváltozik, és egy másik aminosavat kódol. A fehérje funkciója megváltozhat, vagy akár el is veszítheti azt. (Pl. sarlósejtes anémia, ahol a hemoglobin egy aminosava megváltozik.)
- Értelmetlen mutáció (nonsense mutation): A kodon stop kodonná változik, ami a fehérjeszintézis idő előtti befejezését eredményezi. Ez általában egy rövid, nem funkcionális fehérjét eredményez.
- Kereteltolódásos mutáció (frameshift mutation): Inszerció vagy deléció, amely nem három nukleotid többszöröse. Ez eltolja az olvasási keretet, és teljesen megváltoztatja a mutáció utáni összes kodont, ami drámai hatással van a fehérje aminosavsorrendjére.
A sejt számos DNS-javító mechanizmussal rendelkezik, amelyek folyamatosan pásztázzák a genómot a hibák után kutatva, és kijavítják azokat. Ezek közé tartozik a direkt javítás, a bázis-excíziós javítás, a nukleotid-excíziós javítás és a mismatch javítás. Ezek a rendszerek kulcsfontosságúak a genetikai integritás fenntartásában és a betegségek megelőzésében.
Bár a mutációk gyakran károsak, ritka esetekben előnyösek is lehetnek, vagy semlegesek, és hozzájárulnak a genetikai variációhoz. Ez a variáció az evolúció alapvető nyersanyaga, amely lehetővé teszi az élőlények alkalmazkodását a változó környezethez és a fajok diverzitásának kialakulását.
DNS és genetikai örökség

A DNS a genetikai örökség alapja, az a molekula, amely generációról generációra hordozza és továbbítja az élőlények tulajdonságait. Az öröklődés alapelveit először Gregor Mendel osztrák szerzetes fedezte fel a 19. században borsón végzett kísérletei során, jóval a DNS molekuláris szerkezetének ismerete előtt. Mendel törvényei (a uniformitás, a hasadás és a független öröklődés törvénye) a mai napig érvényesek, és a genetikai öröklődés alapkövei.
Minden élőlény genomja a teljes genetikai információt tartalmazza, amely a DNS-ben van kódolva. Az emberi genom körülbelül 3,2 milliárd bázispárból áll, amelyek 23 pár kromoszómán helyezkednek el. Ezek a kromoszómák tartalmazzák a géneket, amelyek a fehérjék szintéziséhez szükséges utasításokat hordozzák, és ezáltal meghatározzák az egyén jellemzőit.
Az öröklődés alapegységei a gének, amelyeknek különböző változatai, az úgynevezett allélek léteznek. Minden ember két allélt örököl minden génből, egyet az anyjától és egyet az apjától. Ezek az allélek lehetnek dominánsak vagy recesszívek. A domináns allélek kifejeződnek, ha csak egy példányban is jelen vannak, míg a recesszív allélek csak akkor fejeződnek ki, ha mindkét allélt recesszív formában örökli az egyén.
A genetikai öröklődés nem korlátozódik kizárólag a sejtmagban lévő DNS-re. Az eukarióta sejtekben a mitokondriumok is tartalmaznak saját, kör alakú DNS-t, az úgynevezett mitokondriális DNS-t (mtDNS). Az mtDNS-t kizárólag az anyától örököljük, mivel a spermiumok mitokondriumai általában nem jutnak be a petesejtbe a megtermékenyítés során. Ez az egyedi öröklődési minta hasznos a populációgenetikai és az evolúciós kutatásokban.
A genetikai örökség megértésében egyre nagyobb szerepet kap az epigenetika. Az epigenetikai változások olyan örökölhető módosulások a génexpresszióban, amelyek nem járnak a DNS-szekvencia megváltozásával. Ezek a módosulások magukban foglalhatják a DNS metilációját vagy a hisztonok kémiai módosításait, amelyek befolyásolják a kromatin szerkezetét és a gének hozzáférhetőségét. Az epigenetika megmagyarázza, hogyan befolyásolhatják a környezeti tényezők, mint például az étrend vagy a stressz, a génjeink működését, és hogyan adódhatnak át ezek a változások a következő generációknak.
A genetikai örökség tanulmányozása alapvető fontosságú a genetikai betegségek diagnosztizálásában és kezelésében. Számos betegség, mint például a cisztás fibrózis, a sarlósejtes anémia vagy a Huntington-kór, egyetlen gén mutációjára vezethető vissza. Más betegségek, mint a cukorbetegség vagy a szívbetegségek, multifaktoriálisak, azaz több gén és környezeti tényező kombinációja okozza őket.
A modern genomiális technológiák, mint a DNS-szekvenálás, lehetővé teszik a teljes genomunk feltérképezését, és mélyebb betekintést nyújtanak az egyéni genetikai örökségbe. Ez forradalmasítja a perszonalizált orvoslást, ahol a kezeléseket az egyén genetikai profiljára szabják, optimalizálva a gyógyszerválasztást és a dózisokat. A genetikai tanácsadás segíti a családokat a genetikai kockázatok megértésében és a tájékozott döntések meghozatalában.
A genetikai örökség nem csupán a szüleinktől kapott tulajdonságok összessége, hanem egy komplex információrendszer, amely folyamatosan kölcsönhatásban áll a környezettel, és formálja azt, akik vagyunk.
A DNS-ismeretek modern alkalmazásai
A DNS szerkezetének és működésének megértése forradalmasította a biológiát és az orvostudományt, megnyitva az utat számos innovatív technológia és alkalmazás előtt. Ezek a felfedezések nem csupán a tudományos kutatást lendítették fel, hanem közvetlen hatással vannak az egészségügyre, a mezőgazdaságra, a bűnüldözésre és még sok más területre.
Genetikai mérnökség és génszerkesztés
A genetikai mérnökség lehetővé teszi a tudósok számára, hogy megváltoztassák az élőlények DNS-ét, új tulajdonságokat adva nekik vagy kijavítva a hibásakat. Ennek legmodernebb eszköze a CRISPR-Cas9 rendszer. Ez a technológia, amely eredetileg a baktériumok immunrendszeréből származik, lehetővé teszi a DNS rendkívül precíz és célzott szerkesztését. A CRISPR képes kivágni a hibás DNS-szakaszokat és helyükre egészségeseket beilleszteni, vagy éppen géneket aktiválni vagy inaktiválni.
A CRISPR-Cas9 óriási potenciállal rendelkezik a genetikai betegségek gyógyításában, mint például a cisztás fibrózis, a sarlósejtes anémia vagy a Huntington-kór. Emellett alkalmazzák a mezőgazdaságban is, ellenállóbb növényfajták létrehozására, és az állattenyésztésben is, a termelési tulajdonságok javítására.
Génterápia
A génterápia célja a betegségek kezelése a hibás gének kijavításával vagy pótlásával. Ez a technológia vírussal (vektorral) juttatja be a korrekciós gént a beteg sejtekbe, ahol az termelni kezdi a hiányzó vagy hibás fehérjét. Már számos génterápiás kezelést engedélyeztek, például bizonyos örökletes szembetegségekre vagy izomsorvadásra.
Bár a génterápia ígéretes, még számos kihívással néz szembe, mint például a génbevitel hatékonysága, az immunválasz elkerülése és a hosszú távú biztonság garantálása. A kutatások azonban folyamatosan zajlanak, és egyre több betegség esetében nyújthat reményt a jövőben.
Forensics és DNS-ujjlenyomat
A DNS-ujjlenyomat (vagy DNS-profilozás) az igazságügyi orvostan alapvető eszköze. A bűncselekmény helyszínén talált biológiai mintákból (vér, haj, nyál, bőrsejtek) kinyert DNS-t elemezve, az egyént nagy pontossággal azonosítani lehet. Ez a technológia forradalmasította a bűnüldözést, segítve a tettesek azonosítását és a tévesen elítéltek felmentését.
A DNS-ujjlenyomat alapja a genomban található ismétlődő, polimorf régiók (STR – Short Tandem Repeats) elemzése, amelyek egyediek minden egyénre nézve (az ikrek kivételével). Ezeket a markereket összehasonlítva lehet megállapítani a minták eredetét.
Perszonalizált orvoslás és farmakogenomika
A perszonalizált orvoslás (vagy precíziós orvoslás) célja, hogy a gyógykezeléseket az egyén genetikai profiljára szabja. A farmakogenomika a gének és a gyógyszerekre adott válasz közötti kapcsolatot vizsgálja. Az egyéni genetikai variációk befolyásolhatják, hogy egy gyógyszer mennyire hatékony vagy milyen mellékhatásokat okoz. A genetikai tesztek segítségével az orvosok előre megjósolhatják, melyik gyógyszer lesz a leghatékonyabb és legbiztonságosabb egy adott beteg számára, optimalizálva a terápiát.
Evolúciós biológia és filogenetika
A DNS-szekvenciák összehasonlítása alapvető eszközzé vált az evolúciós biológia és a filogenetika területén. A fajok közötti genetikai különbségek elemzésével a tudósok rekonstruálni tudják az evolúciós kapcsolatokat, megérteni a fajok eredetét és diverzifikációját, valamint nyomon követni a betegségek terjedését.
A mitokondriális DNS és az Y-kromoszóma DNS-ének elemzése például segített az emberi migrációs minták feltérképezésében és az emberiség közös őseinek azonosításában.
Ezek az alkalmazások csak néhány példát mutatnak be a DNS-ismeretek hatalmas és növekvő hatóköréből. A technológia fejlődésével és a genetikai adatok elemzésének képességével várhatóan még több áttörés várható a jövőben, amelyek tovább formálják az életünket és a világról alkotott képünket.
Etikai és társadalmi megfontolások
A DNS-sel kapcsolatos tudományos és technológiai fejlődés, bár hatalmas lehetőségeket rejt magában, számos etikai és társadalmi kérdést is felvet, amelyekre a társadalomnak közösen kell válaszokat találnia. A genetikai információhoz való hozzáférés, a génszerkesztés morális határai és a genetikai technológiák társadalmi hatásai mind olyan témák, amelyek alapos megfontolást igényelnek.
Genetikai adatvédelem és magánélet
A genetikai adatok rendkívül érzékeny személyes információkat tartalmaznak, amelyek nem csupán az egyénre, hanem annak családtagjaira és jövőbeli generációira is vonatkoznak. A genetikai tesztek terjedésével felmerül a kérdés, hogyan lehet biztosítani ezeknek az adatoknak a védelmét az illetéktelen hozzáférés, a diszkrimináció vagy a visszaélések ellen. Ki férhet hozzá a genetikai információinkhoz? Milyen célra használható fel? Megengedhető-e, hogy biztosítótársaságok vagy munkáltatók felhasználják ezeket az adatokat döntéseik meghozatalakor?
Számos országban már születtek jogszabályok a genetikai diszkrimináció megelőzésére, de a technológia gyors fejlődése miatt a szabályozásnak folyamatosan alkalmazkodnia kell az új kihívásokhoz.
Génszerkesztés és „designer babák”
A génszerkesztési technológiák, mint például a CRISPR-Cas9, lehetővé teszik a DNS precíz módosítását. Bár ez óriási reményt ad a genetikai betegségek gyógyítására, felmerül a kérdés, hol húzódnak a morális határok. Megengedhető-e a génszerkesztés az emberi embriókon a súlyos betegségek megelőzése céljából, ha a változások öröklődnek a következő generációkban?
A legnagyobb etikai dilemma azonban az úgynevezett „designer babák” koncepciójával kapcsolatos. Ha képesek lennénk „javítani” az embriókat, eljuthatunk-e odáig, hogy „tervezett” gyermekeket hozzunk létre, akik bizonyos kívánt tulajdonságokkal (pl. magasabb intelligencia, fizikai erő) rendelkeznek? Ez súlyos társadalmi egyenlőtlenségekhez vezethet, és felveti a fajunk integritásával kapcsolatos kérdéseket.
Hozzáférési egyenlőtlenségek
A modern genetikai technológiák, mint a génterápia vagy a perszonalizált orvoslás, gyakran rendkívül drágák és nem mindenki számára elérhetők. Ez súlyosbíthatja az egészségügyi egyenlőtlenségeket a társadalmon belül és a világ országai között. Hogyan biztosítható, hogy ezek a forradalmi kezelések ne csak a gazdagok kiváltságai legyenek, hanem mindenki számára hozzáférhetővé váljanak, akinek szüksége van rájuk?
A géntechnológia környezeti hatásai
A génmódosított szervezetek (GMO-k) mezőgazdasági alkalmazása is vitatott. Bár a génmódosított növények számos előnnyel járhatnak (pl. nagyobb terméshozam, kártevő-ellenállás), felmerülnek aggályok a környezeti hatásokkal (pl. a vadon élő fajok génjeinek módosulása), az emberi egészségre gyakorolt hosszú távú hatásokkal és a biológiai sokféleségre gyakorolt esetleges negatív hatásokkal kapcsolatban. A tudományos konszenzus szerint a jelenleg forgalomban lévő GMO-k biztonságosak, de a társadalmi elfogadottság továbbra is kihívást jelent.
Ezek az etikai és társadalmi kérdések nem könnyűek, és nem léteznek egyszerű válaszok. A nyílt párbeszéd, a tudósok, etikusok, jogalkotók és a nyilvánosság bevonásával, elengedhetetlen ahhoz, hogy felelősségteljesen kezeljük a DNS-technológiák által kínált lehetőségeket és kihívásokat, biztosítva, hogy az emberiség javát szolgálják, miközben megőrizzük alapvető értékeinket.







