
A sejtek belső kommunikációjának bonyolult hálózatában számos molekula tölt be kulcsfontosságú szerepet, biztosítva a környezeti ingerekre adott precíz és gyors válaszokat. Ezen molekulák közül az egyik legősibb és leguniverzálisabb a ciklikus adenozin-monofoszfát, röviden cAMP. Ez a kis molekula, amelyet az 1950-es évek végén Earl Sutherland fedezett fel, forradalmasította a sejtjelátvitelről alkotott képünket, és a „második hírvivő” koncepciójának alapjává vált. A cAMP nem csupán egy egyszerű közvetítő; komplex szerkezetével és dinamikus szabályozásával a sejtfunkciók széles skáláját befolyásolja, a hormonális válaszoktól kezdve a génexpresszióig, az idegrendszeri plaszticitástól a metabolikus folyamatok finomhangolásáig.
A sejtmembránon található receptorokhoz kötődő elsődleges hírvivők, mint például a hormonok és neurotranszmitterek, ritkán jutnak be a sejt belsejébe. Ehelyett a jelet a sejt belsejébe kell továbbítani, ahol az megfelelő választ vált ki. Itt lép színre a cAMP, amely a sejten belüli jelátviteli kaszkádok egyik legfontosabb elemeként funkcionál. A cAMP koncentrációjának gyors és pontos változásai teszik lehetővé, hogy a sejtek adaptálódjanak a változó környezeti feltételekhez, fenntartsák a homeosztázist, és koordinált módon reagáljanak a külső ingerekre. Megértése elengedhetetlen a sejtfiziológia, a farmakológia és számos betegség patomechanizmusának mélyebb feltárásához.
A ciklikus AMP nem csupán egy molekula, hanem egy ősi és rendkívül adaptív mechanizmus központi eleme, amely a sejtek túlélését és működését biztosítja a legkülönfélébb biológiai rendszerekben.
A jelen cikkben részletesen megvizsgáljuk a cAMP szerkezetét, annak szintézisét és lebontását szabályozó enzimatikus mechanizmusokat, valamint a főbb effektor molekulákat, amelyek közvetítik a cAMP jelét. Különös figyelmet fordítunk a cAMP jelátviteli útvonalak térbeli és időbeli szabályozására, amely elengedhetetlen a specifikus sejtválaszok kialakulásához. Végül áttekintjük a cAMP rendkívül sokrétű szerepét különböző fiziológiai folyamatokban, és betekintést nyerünk abba, hogy a cAMP jelátvitel diszregulációja hogyan járulhat hozzá számos betegség kialakulásához, valamint hogyan hasznosítható ez a tudás a gyógyszerfejlesztésben.
A ciklikus AMP kémiai szerkezete és alapvető tulajdonságai
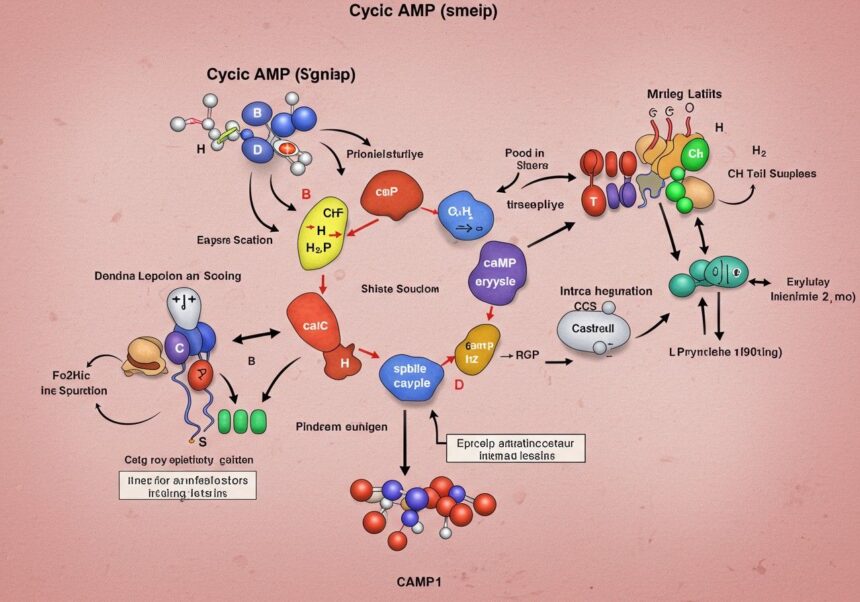
A ciklikus adenozin-monofoszfát (cAMP) kémiai szempontból egy nukleotid származék, amely az adenozin-trifoszfátból (ATP) képződik. Szerkezete az adenin bázisból, egy ribóz cukorból és egy foszfátcsoportból áll. Ami azonban megkülönbözteti a hagyományos AMP-től, az a foszfátcsoport elhelyezkedése: a ribóz gyűrűjének 3′ és 5′ szénatomjához egyaránt kovalensen kötődik, egy belső, ciklikus foszfodiészter kötést hozva létre. Ez a ciklikus szerkezet adja a molekula nevét és egyedi biológiai tulajdonságait.
A foszfodiészter kötés kialakulása egy kondenzációs reakció során megy végbe, melynek során két hidroxilcsoport (a ribóz 3′ és 5′ pozíciójában) reagál a foszfátcsoporttal, vízkilépés közben. Ennek a ciklikus elrendezésnek köszönhetően a cAMP egy viszonylag stabil molekula, ám mégis képes gyorsan lebomlani, biztosítva ezzel a jelátviteli rendszer dinamikus szabályozását. A ciklikus foszfátgyűrű elektrosztatikus tulajdonságai és a molekula térbeli konformációja kulcsfontosságúak az effektor fehérjékkel való specifikus kölcsönhatásban.
A cAMP molekula vízoldékony, ami lehetővé teszi számára, hogy szabadon diffundáljon a citoplazmában, és eljusson a különböző sejtszervecskékhez és effektor fehérjékhez. Ez a tulajdonság alapvető fontosságú a jel gyors és hatékony terjesztéséhez a sejt belsejében. A molekula kis mérete és relatív stabilitása ideális „második hírvivővé” teszi, amely képes a sejtfelszíni receptorokhoz érkező jeleket amplifikálni és továbbítani a sejtmagba vagy más célpontokhoz.
Összehasonlítva más ciklikus nukleotidokkal, mint például a ciklikus guanozin-monofoszfáttal (cGMP), a cAMP szerkezeti hasonlóságokat mutat, de a bázis (adenin vs. guanin) különbsége miatt eltérő effektor fehérjékhez kötődik és különböző jelátviteli útvonalakat aktivál. Bár vannak átfedések és keresztreakciók, a legtöbb esetben a cAMP és a cGMP specifikus utakat aktivál, lehetővé téve a sejtek számára, hogy differenciált válaszokat adjanak a különböző ingerekre.
A ciklikus AMP szintézise: az adenilát-cikláz enzim
A cAMP szintézisének központi enzime az adenilát-cikláz (AC), egy transzmembrán fehérje, amely a sejtmembrán belső oldalán található, vagy egyes izoformák esetében a citoszolban. Ez az enzim katalizálja az adenozin-trifoszfát (ATP) molekula ciklikus AMP-vé és pirofoszfáttá történő átalakítását. A reakció során az ATP két foszfátcsoportja (a béta és gamma foszfátok) hasad le, és a maradék alfa foszfát egy ciklikus kötést hoz létre a ribóz 3′ és 5′ hidroxilcsoportjaival.
Az adenilát-cikláz aktivitását szigorúan szabályozzák, elsősorban a G-fehérjék családjába tartozó heterotrimer G-fehérjék. A G-fehérjék a G-fehérje-kapcsolt receptorok (GPCR-ek) downstream effektorai, és kulcsszerepet játszanak a sejtfelszíni jelek intracelluláris továbbításában. Amikor egy ligand (pl. hormon, neurotranszmitter) kötődik egy GPCR-hez, az aktiválja a hozzá kapcsolódó G-fehérjét, amely GTP-t köt, és alegységeire disszociál.
A G-fehérjék szerepe az adenilát-cikláz szabályozásában
A G-fehérjék többféle módon befolyásolhatják az adenilát-cikláz aktivitását:
- Stimuláló G-fehérjék (Gs): A Gs alegysége, a Gsα, GTP kötése után disszociál a Gβγ alegységtől, és közvetlenül aktiválja az adenilát-ciklázt. Ez a cAMP termelődésének fokozódásához vezet. Számos hormon, mint például az adrenalin (béta-adrenerg receptorokon keresztül), a glukagon, a TSH és az ACTH, a Gs útvonalon keresztül fejti ki hatását.
- Inhibitor G-fehérjék (Gi): A Gi alegysége, a Giα, szintén GTP kötése után disszociál, de ellentétben a Gsα-val, gátolja az adenilát-cikláz aktivitását. Ezáltal csökken a cAMP szintje a sejtben. Például a szomatosztatin és egyes dopamin receptorok a Gi útvonalon keresztül hatnak.
- Egyéb G-fehérjék (Gq, G12/13): Bár ezek elsősorban más effektorokat (pl. foszfolipáz C) szabályoznak, a Gβγ alegységük is modulálhatja az adenilát-cikláz egyes izoformáit, hol stimuláló, hol gátló hatást kifejtve.
Az adenilát-cikláz család legalább kilenc membránkötött izoformát (AC1-AC9) és egy oldható izoformát (sAC) foglal magában, amelyek mindegyike eltérő szöveti eloszlással és szabályozási mechanizmusokkal rendelkezik. Ez a diverzitás biztosítja, hogy a különböző sejtek és szövetek specifikusan reagálhassanak a külső ingerekre, és finomhangolják a cAMP szintjét a specifikus fiziológiai igényekhez.
Például, az AC1 és AC8 izoformák kalcium/kalmodulin-függő módon aktiválódnak, ami egy fontos kapcsolódási pontot jelent a cAMP és a kalcium jelátviteli rendszerek között. Az AC5 és AC6 viszont kalcium által gátolható, ami a szívizomban különösen fontos a cAMP szint szabályozásában. Az sAC (oldható adenilát-cikláz) nem G-fehérjék által szabályozott, hanem közvetlenül a bikarbonát és a kalcium ionok koncentrációjára érzékeny, és a sejt belső környezeti változásaira reagál, például a spermiumokban játszik kulcsszerepet azok motilitásának szabályozásában.
Az adenilát-cikláz aktivitásának precíz szabályozása tehát alapvető a sejt homeosztázisának fenntartásához és a környezeti változásokra adott adaptív válaszokhoz. Bármilyen zavar a G-fehérje-AC interakciókban súlyos következményekkel járhat, és számos betegség patogenezisében szerepet játszhat.
A ciklikus AMP lebontása: a foszfodiészterázok (PDE-k)
Ahhoz, hogy a cAMP jelátviteli rendszer dinamikus és gyorsan reverzibilis legyen, a cAMP szintézise mellett annak lebontása is kulcsfontosságú. A ciklikus nukleotid foszfodiészterázok (PDE-k) enzimek felelősek a cAMP (és gyakran a cGMP) hidrolíziséért, azaz a ciklikus foszfodiészter kötés elhasításáért, ami inaktív 5′-AMP-t eredményez. Ez a lebontási folyamat biztosítja a cAMP jel gyors kikapcsolását, lehetővé téve a sejt számára, hogy készen álljon a következő jel vételére.
A PDE enzimek egy nagy és heterogén családot alkotnak, amelyek legalább 11 fő családot (PDE1-PDE11) foglalnak magukban, és ezeken belül számos izoenzim létezik. Ezek az izoenzimek eltérő szubsztrát-specifitással (cAMP-specifikus, cGMP-specifikus vagy kettős specifitású), eltérő szöveti eloszlással, intracelluláris lokalizációval és szabályozási mechanizmusokkal rendelkeznek. Ez a diverzitás teszi lehetővé a cAMP jelátvitel rendkívül finomhangolt szabályozását különböző sejttípusokban és sejtszervecskékben.
A foszfodiészteráz enzimek főbb családjai és jellemzőik
A PDE családok közül néhány kiemelten fontos a cAMP metabolizmus szempontjából:
| PDE család | Fő szubsztrát | Jellemzők és szerep |
|---|---|---|
| PDE1 | cAMP, cGMP | Kalcium/kalmodulin-függő aktiváció. Szerepe van az érrendszerben, idegrendszerben. |
| PDE2 | cAMP, cGMP | cGMP által aktiválódik, ami gátolja a cAMP lebontását. Fontos a keresztreakciókban. |
| PDE3 | cAMP >> cGMP | Erősen cAMP-specifikus. Szerepe a szívizomban (pozitív inotróp hatás), vérlemezkékben. Gátlása pl. cilostazollal. |
| PDE4 | cAMP-specifikus | A legfontosabb cAMP-specifikus PDE család. Négy alcsalád (A, B, C, D) és számos izoforma. Kiemelt szerepe van a gyulladásos folyamatokban, immunsejtekben, idegrendszerben. Gátlása pl. roflumilaszt. |
| PDE7 | cAMP-specifikus | Immunsejtekben, T-limfocitákban expresszálódik, szerepe van az immunválasz szabályozásában. |
| PDE8 | cAMP-specifikus | Spermiumokban, immunsejtekben található. |
| PDE10 | cAMP, cGMP | Kiemelten expresszálódik az agy striátumában, potenciális terápiás célpont neurológiai betegségekben. |
| PDE11 | cAMP, cGMP | Hím reproduktív szervekben, agyban, vese sejtjeiben. |
A PDE-k aktivitását nemcsak a szubsztrátkoncentráció, hanem számos más mechanizmus is szabályozza, beleértve a foszforilációt (pl. PKA vagy PKG által), a kalcium/kalmodulin kötődést, valamint a fehérje-fehérje kölcsönhatásokat. Ez a komplex szabályozás lehetővé teszi, hogy a cAMP szintje precízen modulálható legyen a sejt különböző kompartmentjeiben, létrehozva lokális cAMP grádienseket, amelyek specifikus sejtválaszokat irányítanak.
A foszfodiészterázok nem csupán kikapcsolják a cAMP jelet, hanem aktívan formálják annak térbeli és időbeli profilját, lehetővé téve a sejt számára a rendkívül differenciált válaszokat.
A PDE-k farmakológiai gátlása rendkívül fontos terápiás stratégia. A PDE gátlók célzottan emelik a cAMP (vagy cGMP) szintjét a sejtben, és így befolyásolják a downstream jelátviteli utakat. Például a koffein és a teofillin nem szelektív PDE gátlók, amelyek stimuláló hatásukat részben a cAMP szint emelésén keresztül fejtik ki. Specifikusabb gátlók, mint a cilostazol (PDE3 gátló) az erek tágítására és a vérlemezkék aggregációjának gátlására szolgál. A sildenafil (Viagra), egy PDE5 gátló, a cGMP lebontását gátolja, ami az erek ellazulásához vezet. A roflumilaszt (PDE4 gátló) a krónikus obstruktív tüdőbetegség (COPD) kezelésére használják gyulladáscsökkentő hatása miatt.
A PDE-k funkciójának megértése és a szelektív gátlók fejlesztése kritikus fontosságú a modern gyógyszerkutatásban, mivel ezek az enzimek potenciális célpontokat jelentenek számos betegség, köztük a gyulladásos betegségek, szív- és érrendszeri betegségek, neurológiai rendellenességek és rák kezelésére.
A cAMP főbb effektor molekulái
A cAMP, mint másodlagos hírvivő, önmagában nem fejti ki a biológiai hatásait, hanem specifikus effektor fehérjékhez kötődve aktiválja vagy modulálja azok működését. A cAMP főbb effektor molekulái közé tartozik a protein kináz A (PKA), az Epac (Exchange Protein Activated by cAMP) fehérjék és a ciklikus nukleotid-függő ioncsatornák (CNG-csatornák). Ezek az effektorok a cAMP koncentrációjának változásaira reagálva downstream jelátviteli kaszkádokat indítanak el, amelyek végső soron a sejt fiziológiai válaszait eredményezik.
A protein kináz A (PKA): a cAMP elsődleges közvetítője
A protein kináz A (PKA), más néven cAMP-függő protein kináz, a cAMP legismertebb és legszélesebb körben tanulmányozott effektorja. Szinte minden eukarióta sejtben jelen van, és a sejt számos alapvető funkcióját szabályozza. A PKA egy heterotetramer enzim, amely két regulatórikus (R) és két katalitikus (C) alegységből áll inaktív állapotban. Az R alegységek tartalmazzák a cAMP kötőhelyeit, míg a C alegységek felelősek a szubsztrátfehérjék szerin és treonin oldalláncainak foszforilációjáért.
A PKA aktivációjának mechanizmusa
Amikor a cAMP szintje emelkedik a sejtben, a cAMP molekulák a PKA regulatórikus alegységeihez kötődnek. Minden R alegység két cAMP kötőhelyet tartalmaz. A cAMP kötődése konformációváltozást idéz elő az R alegységeken, ami csökkenti azok affinitását a C alegységekhez. Ennek következtében a két katalitikus alegység disszociál a regulatórikus alegységekről, és aktívvá válik. Az aktív C alegységek ezután számos intracelluláris fehérjét foszforilálnak, megváltoztatva azok aktivitását, lokalizációját vagy kölcsönhatásait más fehérjékkel.
A PKA által foszforilált célfehérjék rendkívül sokrétűek, és magukban foglalják a transzkripciós faktorokat, metabolikus enzimeket, ioncsatornákat, transzportereket, és strukturális fehérjéket. Példák a PKA célpontjaira:
- CREB (cAMP Response Element-Binding protein): Egy transzkripciós faktor, amely foszforilációja után a génpromóterek cAMP-válasz elemeihez (CRE) kötődik, és aktiválja a génexpressziót. Ez kulcsszerepet játszik a hosszú távú memória konszolidációjában és a génszabályozásban.
- Glikogén foszforiláz kináz és glikogén szintáz: A PKA foszforilálja és aktiválja a glikogén foszforiláz kinázt, ami a glikogén lebontásához vezet. Ezzel egyidejűleg foszforilálja és inaktiválja a glikogén szintázt, gátolva a glikogén szintézist. Ez a mechanizmus biztosítja, hogy stresszhelyzetben (pl. adrenalin hatására) gyorsan mobilizálódjon a glükóz.
- Ioncsatornák: A PKA foszforilálhatja például a szívizomsejtekben található kalciumcsatornákat (L-típusú Ca2+ csatornák), növelve a kalcium beáramlását és ezzel a szívizom kontraktilitását.
- Vízcsatornák (aquaporinok): A vese gyűjtőcsatornáiban a PKA foszforilálja az aquaporin-2-t, elősegítve annak beépülését a sejtmembránba, és növelve a víz reabszorpcióját az antidiuretikus hormon (ADH) hatására.
A PKA által közvetített jelek diverzitását az is biztosítja, hogy a PKA nem diffundál szabadon a citoplazmában, hanem specifikus AKAP (A-Kinase Anchoring Proteins) fehérjékhez kötődik. Az AKAP-ok a PKA-t a sejt specifikus részeire (pl. membrán, citoszkeleton, sejtmag) horgonyozzák, és jelátviteli komplexekbe szervezik más enzimekkel és szubsztrátokkal. Ez a kompartmentalizáció biztosítja, hogy a cAMP jel lokalizált és specifikus válaszokat váltson ki, elkerülve a nem kívánt, általános foszforilációt.
Az Epac (Exchange Protein Activated by cAMP) fehérjék
Az Epac fehérjék (Epac1 és Epac2) a cAMP egy másik fontos effektor családját képviselik, amelyeket az 1990-es évek végén fedeztek fel. Az Epac fehérjék GTPáz-aktiváló faktorok (GEF-ek) a kis GTPázok, különösen a Rap1 és Rap2 számára. A Rap1 és Rap2 a Ras szupercsaládba tartozó kis GTPázok, amelyek számos sejtfunkcióban, például a sejtadhézióban, szekrécióban, sejtpolaritásban és génexpresszióban játszanak szerepet.
Az Epac aktivációjának mechanizmusa és szerepe
A cAMP kötődése az Epac fehérjék regulatórikus doménjéhez konformációváltozást idéz elő, ami felszabadítja az Epac GEF-doménjének gátlását. Az aktív Epac ezután elősegíti a Rap1 és Rap2 GDP-ről GTP-re való cseréjét, ezáltal aktiválva ezeket a kis GTPázokat. Az aktivált Rap1 és Rap2 továbbítják a jelet downstream effektorokhoz, mint például a Raf-1, B-Raf vagy a PLCε.
Az Epac útvonal sokféle sejtfunkciót szabályoz, gyakran párhuzamosan vagy együttműködve a PKA útvonallal. Néhány példa az Epac szerepére:
- Sejtadhézió és migráció: Az Epac/Rap1 útvonal kulcsszerepet játszik az integrinek aktiválásában és a sejt-sejt, valamint sejt-extracelluláris mátrix adhézió szabályozásában, befolyásolva a sejtek mozgását és morfológiáját.
- Inzulin szekréció: A béta-sejtekben az Epac2 fokozza a glükóz-indukált inzulin szekréciót a kalcium jelátvitel modulálásán keresztül.
- Neurotranszmitter felszabadulás: Az idegsejtekben az Epac szerepet játszik a szinaptikus plaszticitásban és a neurotranszmitterek exocitózisában.
- Endotél barrier integritása: Az Epac/Rap1 útvonal erősíti az endotél sejtek közötti tight junction-öket, ami fontos az érpermeabilitás szabályozásában.
Bár a PKA és az Epac is cAMP-re reagál, gyakran eltérő módon járulnak hozzá a sejtválaszokhoz, és specifikus, vagy éppen kooperatív hatásokat fejtenek ki. A kutatások egyre inkább rávilágítanak arra, hogy a cAMP jelátvitel finomhangolása a PKA és Epac útvonalak közötti egyensúlyon múlik.
Ciklikus nukleotid-függő ioncsatornák (CNG-csatornák)
A ciklikus nukleotid-függő ioncsatornák (CNG-csatornák) egy harmadik fontos effektor osztályt képviselnek, amelyek közvetlenül a cAMP (vagy cGMP) kötődésére reagálnak anélkül, hogy kináz aktivitásra lenne szükség. Ezek a csatornák ligand-kapcsolt ioncsatornák, amelyek a cAMP közvetlen kötődése hatására nyílnak meg, lehetővé téve a kationok, főleg a nátrium és kalcium ionok áramlását a sejtmembránon keresztül.
A CNG-csatornák szerkezete és működése
A CNG-csatornák tetramer fehérjék, amelyek minden alegysége egy ciklikus nukleotid kötőhelyet tartalmaz. A cAMP (vagy cGMP) kötődése konformációváltozást indukál a csatornában, ami a pórus megnyílásához vezet. Ezek a csatornák különösen fontosak az érzékszervekben, ahol a gyors és közvetlen ionáramlás elengedhetetlen a szenzoros transzdukcióhoz.
Példák a CNG-csatornák szerepére:
- Fototranszdukció: A retina pálcika és csapsejtjeiben a cGMP-függő ioncsatornák (CNG-csatornák) kulcsszerepet játszanak a fényérzékelésben. Sötétben magas a cGMP szint, ami nyitva tartja a csatornákat. Fény hatására a cGMP lebomlik, a csatornák záródnak, ami hiperpolarizációt okoz, és elindítja a vizuális jelet.
- Szaglás: Az orrnyálkahártya szaglóreceptor sejtjeiben a cAMP-függő ioncsatornák (CNG-csatornák) felelősek a szagingerek érzékeléséért. A szaganyagok kötődése a receptorokhoz cAMP termelést indukál, ami megnyitja a CNG-csatornákat, depolarizálva a sejtet és elindítva a szaglási jelet.
- Kardiális funkciók: A szívben is találhatók HCN (Hyperpolarization-activated Cyclic Nucleotide-gated) csatornák, amelyek funkciója hasonló a CNG-csatornákéhoz, és a szívritmus szabályozásában játszanak szerepet a cAMP modulációján keresztül.
Összefoglalva, a cAMP jelátvitel komplexitása abban rejlik, hogy számos különböző effektor molekulán keresztül képes hatását kifejteni. A PKA, az Epac és a CNG-csatornák eltérő mechanizmusokon keresztül, de együttesen biztosítják a sejt számára a rendkívül sokoldalú és finomhangolt válaszokat a cAMP szintjének változásaira.
A cAMP jelátviteli útvonalak térbeli és időbeli szabályozása
A cAMP jelátviteli rendszer nem csupán egy egyszerű, diffúz folyamat, ahol a cAMP egyenletesen eloszlik a citoplazmában és aktiválja az összes elérhető effektor molekulát. Éppen ellenkezőleg, a cAMP jelátvitel térbeli és időbeli szabályozása kulcsfontosságú a specifikus és precíz sejtválaszok kialakításában. A sejtek képesek lokális cAMP mikrodoméneket létrehozni, fenntartani és lebontani, amelyek lehetővé teszik a különböző ingerekre adott differenciált válaszokat még ugyanazon a sejten belül is.
Az AKAP (A-Kinase Anchoring Proteins) fehérjék szerepe
Az egyik legfontosabb mechanizmus, amely a cAMP jelátvitel térbeli szerveződését biztosítja, az AKAP (A-Kinase Anchoring Proteins) fehérjék rendszere. Az AKAP-ok egy heterogén fehérjecsaládot alkotnak, amelyek képesek a PKA regulatórikus alegységeihez kötődni, és ezáltal a kinázt specifikus intracelluláris helyekre horgonyozni. Ezen túlmenően, sok AKAP nem csupán PKA-t köt, hanem más jelátviteli molekulákat is, mint például adenilát-ciklázokat, foszfodiészterázokat (PDE-ket), foszfatázokat, vagy más kinázokat, létrehozva így egy multifunkcionális jelátviteli komplexet, vagy más néven szignaloszómát.
Az AKAP-ok szerepe többszörös:
- Lokalizáció: Az AKAP-ok biztosítják a PKA és más kapcsolódó enzimek specifikus lokalizációját a sejt különböző kompartmentjeiben, például a plazmamembránban, a mitokondriumban, az endoplazmatikus retikulumban vagy a sejtmagban. Ezáltal a cAMP által aktivált PKA csak a közelében lévő, specifikus szubsztrátokat foszforilálja.
- Jelátviteli hatékonyság: Azáltal, hogy a cAMP termelő, lebontó és effektor enzimeket fizikailag közel hozzák egymáshoz, az AKAP-ok növelik a jelátvitel hatékonyságát és sebességét. A cAMP koncentrációjának változásai lokálisan, gyorsan és célzottan képesek befolyásolni a PKA aktivitását.
- Jel specifikussága: Az AKAP-ok biztosítják a jel specifikusságát. Egy adott receptor aktiválódása lokális cAMP emelkedést okozhat, amely csak a megfelelő AKAP-hoz kötött PKA-t aktiválja, és csak az adott jelátviteli komplexhez tartozó célfehérjéket foszforilálja. Ez megakadályozza a jel „szétkenődését” és a nem kívánt mellékhatásokat.
- Jel integráció: Az AKAP-ok gyakran integrálják a különböző jelátviteli útvonalakat. Például egy AKAP komplex tartalmazhat PKA-t és protein kináz C-t (PKC) is, lehetővé téve a cAMP és a kalcium/diacilglicerol útvonalak közötti kereszthatásokat és finomhangolt válaszokat.
Például, a szívizomsejtekben az AKAP79/150 egy fontos AKAP, amely a plazmamembránhoz horgonyozza a PKA-t, és a béta-adrenerg receptorok által kiváltott cAMP jelet a Ca2+ csatornákhoz irányítja, fokozva a szívizom összehúzódását. Az AKAP18δ a szívizom szarkoplazmás retikulumához kötődik, és a SERCA2a kalcium pumpa aktivitását modulálja a PKA foszforilációján keresztül.
Mikrodomének és lokalizált cAMP jelátvitel
Az AKAP-okon túlmenően, a sejtek képesek cAMP mikrodoméneket létrehozni, amelyek a cAMP koncentrációjának lokális ingadozásait jelentik. Ezek a mikrodomének a sejtmembrán specifikus régióiban, például lipid tutajokban (lipid rafts) vagy a plazmamembrán és a szarkoplazmás retikulum közötti szoros kapcsolódási pontokon alakulhatnak ki. Ezekben a mikrodoménekben az adenilát-ciklázok, PDE-k és effektor molekulák (PKA, Epac, CNG-csatornák) térben szervezetten helyezkednek el.
A PDE-k kulcsszerepet játszanak a cAMP mikrodomének fenntartásában azáltal, hogy gyorsan lebontják a cAMP-t a szintézis helyétől távolabb eső területeken. Ez létrehoz egy cAMP grádienst, ahol a cAMP koncentrációja magasabb a szintézis helyén (pl. egy aktivált adenilát-cikláz közelében) és alacsonyabb a környező területeken. Ez a grádiens lehetővé teszi, hogy egy adott receptor aktiválódása csak a közvetlen közelében lévő effektorokat aktiválja, míg a sejt más részein a cAMP szint változatlan marad.
A térbeli grádiensek és a lokalizált cAMP jelátvitel elengedhetetlen a sejt polaritásának, a sejtek migrációjának, a szinaptikus plaszticitásnak és számos más folyamatnak a szabályozásához. Például az immunsejtekben a kemotaktikus ingerek lokális cAMP grádienseket hoznak létre, amelyek irányítják a sejtek mozgását a gyulladás helyére.
Időbeli dinamika és pulzáló cAMP jelek
A térbeli szabályozás mellett a cAMP jelátvitel időbeli dinamikája is rendkívül fontos. A cAMP szintje nem statikus, hanem gyorsan emelkedhet és csökkenhet, gyakran pulzáló vagy oszcilláló mintázatot mutatva. Ezek az időbeli mintázatok is hordozhatnak információt a sejt számára, és eltérő választ válthatnak ki, mint egy tartósan magas cAMP szint.
Az időbeli dinamikát az adenilát-ciklázok és PDE-k aktivitásának összehangolt szabályozása határozza meg. A gyors aktiválódás és inaktiválódás, valamint a visszacsatolási mechanizmusok (pl. PKA által mediált foszforilációja az AC-nek vagy a PDE-nek) hozzájárulnak a cAMP oszcillációk kialakulásához. Ezek az oszcillációk különösen fontosak lehetnek a sejtosztódás, a differenciáció és a génexpresszió szabályozásában, ahol a jel időzítése kritikus.
Összefoglalva, a cAMP jelátvitel messze túlmutat egy egyszerű „be-ki” kapcsolón. A kompartmentalizáció, az AKAP-ok, a mikrodomének és az időbeli dinamika révén a sejt képes rendkívül komplex és finomhangolt módon értelmezni a cAMP jeleket, és differenciált válaszokat adni a környezeti ingerekre. Ezen mechanizmusok megértése alapvető a sejtfunkciók és a betegségek molekuláris alapjainak feltárásához.
A cAMP szerepe különböző fiziológiai folyamatokban
A ciklikus AMP rendkívül sokoldalú molekula, amely szinte minden eukarióta sejtben jelen van, és a sejtfunkciók széles skáláját szabályozza. Jelátviteli útvonala számos életfontosságú folyamatban kulcsszerepet játszik, a hormonális válaszoktól az idegrendszeri működésig, a metabolikus szabályozástól az immunválaszig.
Hormonális válaszok és metabolikus reguláció
A cAMP a hormonális jelátvitel egyik legismertebb és legfontosabb közvetítője. Számos hormon, amelyek a G-fehérje-kapcsolt receptorokhoz (GPCR-ekhez) kötődnek, hatásukat a cAMP szintjének modulálásán keresztül fejtik ki. Ezek a hormonok befolyásolják a sejt metabolizmusát, növekedését és differenciációját.
- Adrenalin (epinefrin) és noradrenalin: A béta-adrenerg receptorokhoz kötődve aktiválják a Gs fehérjét és az adenilát-ciklázt, ami cAMP emelkedéshez vezet. Ez a szívizomban pozitív inotróp és kronotróp hatást eredményez (erősebb és gyorsabb szívverés), a májban és izmokban fokozza a glikogén lebontását és a glükóz felszabadulását (glikogenolízis), a zsírsejtekben pedig a zsírbontást (lipolízis). Ez a „harcolj vagy menekülj” válasz alapja.
- Glukagon: A hasnyálmirigy alfa-sejtjei által termelt hormon, amely a máj glukagon receptoraihoz kötődve fokozza a cAMP termelést. Ez serkenti a glikogenolízist és a glükoneogenezist, emelve a vércukorszintet.
- Pajzsmirigy-stimuláló hormon (TSH): A TSH a pajzsmirigy sejtjeiben cAMP emelkedést okoz, ami serkenti a pajzsmirigyhormonok (T3 és T4) szintézisét és felszabadulását.
- Adrenokortikotrop hormon (ACTH): Az ACTH a mellékvesekéreg sejtjeiben cAMP-függő útvonalon keresztül stimulálja a kortizol és más szteroid hormonok szintézisét.
- Vazopresszin (ADH): A vese gyűjtőcsatornáinak sejtjeiben a vazopresszin cAMP emelkedést okoz, ami a PKA aktiválásán keresztül az aquaporin-2 vízcsatornák membránba való beépülését segíti elő, növelve a víz reabszorpcióját és szabályozva a vízháztartást.
Ezeken kívül a cAMP a zsíranyagcsere (lipolízis), a koleszterin szintézis, és az inzulin szekréció szabályozásában is részt vesz, így alapvető fontosságú a szervezet metabolikus homeosztázisának fenntartásában.
Idegrendszeri funkciók és szinaptikus plaszticitás
Az idegrendszerben a cAMP jelátvitel kritikus szerepet játszik a neurotranszmisszióban, a szinaptikus plaszticitásban, a memória konszolidációjában és a tanulási folyamatokban. Számos neurotranszmitter (pl. dopamin, szerotonin, noradrenalin) hatását cAMP-függő útvonalakon keresztül fejti ki.
- Szinaptikus plaszticitás: A hosszú távú potenciáció (LTP) és a hosszú távú depresszió (LTD), amelyek a memória és a tanulás alapját képező szinaptikus erősség változásai, gyakran magukban foglalják a cAMP/PKA útvonal aktiválódását. A PKA foszforilálja az ioncsatornákat és a transzkripciós faktorokat (pl. CREB), amelyek módosítják a szinapszisok szerkezetét és működését, valamint a génexpressziót.
- Neurotranszmitter felszabadulás: A cAMP modulálja a preszinaptikus terminálisokban a neurotranszmitterek felszabadulását, befolyásolva a szinaptikus átvitelt.
- Neuronális fejlődés és túlélés: A cAMP fontos szerepet játszik a neuronok növekedésében, differenciálódásában és túlélésében, különösen a neurotróf faktorok által kiváltott jelátviteli útvonalakon keresztül.
Az agyban a cAMP szintek diszregulációja számos neurológiai és pszichiátriai betegséggel hozható összefüggésbe, mint például a depresszió, szorongás, skizofrénia, Parkinson-kór és Alzheimer-kór.
Szív- és érrendszer
A cAMP jelátvitel alapvető a szív- és érrendszer működésének szabályozásában. A béta-adrenerg receptorok aktiválódása a szívizomban cAMP szint emelkedést okoz, ami a PKA aktiválásán keresztül foszforilálja a kalciumcsatornákat, a foszfolambánt (SR Ca2+-ATPáz szabályozója) és a troponin I-t. Ez a foszforiláció növeli a szívizom kontrakciós erejét (pozitív inotróp hatás) és a relaxáció sebességét (pozitív lusitróp hatás), valamint a szívfrekvenciát (pozitív kronotróp hatás).
Az erek simaizomsejtjeiben a cAMP emelkedés relaxációt és értágulatot okoz, mivel a PKA foszforilálja a miozin könnyűlánc kinázát (MLCK), gátolva annak aktivitását, és ezáltal a miozin fej foszforilációját, ami a kontrakcióhoz szükséges. Ez a mechanizmus fontos a vérnyomás szabályozásában.
Immunrendszer és gyulladás
Az immunrendszerben a cAMP általában gyulladáscsökkentő és immunszuppresszív hatású. A cAMP szint emelkedése a különböző immunsejtekben (pl. limfociták, makrofágok, neutrofilek) gátolja azok aktiválódását, proliferációját és citokin termelését. Ez a mechanizmus hozzájárul a gyulladásos válasz modulálásához és a túlzott immunreakciók megakadályozásához.
- Gyulladáscsökkentő hatás: A PKA foszforilálhatja a nukleáris faktor kappa B (NF-κB) jelátviteli útvonal komponenseit, gátolva a gyulladásos gének expresszióját.
- Asztma és COPD: A hörgők simaizomsejtjeiben a cAMP emelkedése bronchodilatációt okoz, ami enyhíti az asztmás rohamokat. A PDE4 gátlók (pl. roflumilaszt) gyulladáscsökkentő hatásuk miatt használatosak krónikus obstruktív tüdőbetegségben (COPD).
Sejtproliferáció és differenciáció
A cAMP jelátvitel komplex módon befolyásolja a sejtproliferációt és differenciációt, a sejttípustól és a kontextustól függően. Egyes sejtekben a cAMP gátolja a proliferációt és elősegíti a differenciációt, míg más sejtekben serkenti a növekedést. Például, a cAMP a Schwann-sejtek differenciációjának és a mielinhüvely képződésének fontos szabályozója. A rák kutatásban is kiemelt figyelmet kap a cAMP szerepe a tumorsejt-növekedés és apoptózis modulálásában.
Ez a felsorolás is jól mutatja, hogy a cAMP jelátviteli rendszer milyen alapvető és sokrétű szerepet játszik a normális sejtműködésben és a szervezet homeosztázisának fenntartásában. Ennek a rendszernek a diszfunkciója számos patofiziológiai állapot kialakulásához vezethet.
A cAMP jelátvitel patofiziológiai vonatkozásai és betegségek
A ciklikus AMP jelátviteli útvonalak diszregulációja számos betegség patogenezisében kulcsszerepet játszik. A cAMP szintjének kóros emelkedése vagy csökkenése, az effektor fehérjék működési zavarai, vagy a jelátviteli komplexek hibás szerveződése mind hozzájárulhatnak különböző klinikai állapotok kialakulásához. A cAMP útvonalak megértése ezért nemcsak a betegségek molekuláris alapjainak feltárásához, hanem új terápiás stratégiák kidolgozásához is elengedhetetlen.
Rák és sejtproliferáció
A cAMP jelátvitel szerepe a rákban komplex és kontextusfüggő. Hagyományosan a cAMP-t tumorellenes hatású molekulaként tartották számon, amely gátolja a sejtproliferációt és elősegíti a differenciációt vagy az apoptózist (programozott sejthalál). Ez a hatás gyakran a PKA-n keresztül valósul meg, amely foszforilálja a sejtciklust szabályozó fehérjéket, vagy aktiválja a tumor szuppresszor gének transzkripcióját.
Azonban a kutatások kimutatták, hogy bizonyos ráktípusokban a cAMP jelátvitel akár tumorpromotáló hatással is bírhat, serkentve a sejtproliferációt, a túlélést és a metasztázist. Ez a kettős szerep a cAMP izoformáinak és effektorainak (PKA, Epac) eltérő expressziójából, lokalizációjából és kölcsönhatásaiból adódhat a különböző daganattípusokban. Például, bizonyos emlőrák sejtekben a megnövekedett cAMP szint fokozhatja a növekedést. A cAMP jelátvitel modulálása ígéretes terápiás célpontot jelenthet a rákellenes kezelésekben, de a specifikus útvonalak és a sejttípus-függő hatások mélyebb megértésére van szükség.
Cukorbetegség és metabolikus szindróma
A cAMP jelátvitel alapvető fontosságú a glükóz-homeosztázis szabályozásában, különösen az inzulin szekréció és az inzulinérzékenység tekintetében. A hasnyálmirigy béta-sejtjeiben a glükóz által kiváltott inzulin szekréciót a cAMP szint emelkedése fokozza, mind a PKA, mind az Epac útvonalakon keresztül. Az Epac2 különösen fontosnak bizonyult a glükóz-érzékelés és az inzulin felszabadulás mediálásában.
A 2-es típusú cukorbetegségben a béta-sejtek működési zavara és az inzulinrezisztencia jellemző. A cAMP jelátvitel hibái, mint például az adenilát-cikláz aktivitásának csökkenése vagy a PDE-k túlműködése, hozzájárulhatnak az inzulin szekréció romlásához. A PDE3 gátlók, amelyek emelik a cAMP szintet, már vizsgálták, mint potenciális gyógyszereket a cukorbetegség kezelésében az inzulin szekréció fokozására.
Szív- és érrendszeri betegségek
A szív- és érrendszeri betegségek, mint a szívelégtelenség, aritmiák és hipertónia, gyakran járnak a cAMP jelátvitel diszregulációjával. A béta-adrenerg receptorok túlzott vagy krónikus stimulációja szívelégtelenséghez vezethet, mivel a tartósan magas cAMP szint és PKA aktiváció hosszú távon károsítja a szívizomsejteket, és rontja a kalcium-homeosztázist.
A PDE-k, különösen a PDE3 és PDE4, kulcsfontosságúak a cAMP lebontásában a szívben. A PDE3 gátlók, mint a milrinon, pozitív inotróp hatásúak és szívelégtelenségben alkalmazhatók, de hosszú távú használatuk korlátozott a mellékhatások, például az aritmiák fokozott kockázata miatt. A PDE-k szelektív gátlása új terápiás lehetőségeket kínálhat a szív- és érrendszeri betegségek kezelésében, a mellékhatások minimalizálásával.
Neurológiai és pszichiátriai rendellenességek
Az agyban a cAMP jelátvitel diszregulációja számos neurológiai és pszichiátriai betegséggel hozható összefüggésbe. A depresszió, szorongás, skizofrénia, bipoláris zavar, Parkinson-kór és Alzheimer-kór mindegyikében találtak bizonyítékot a cAMP/PKA/CREB útvonalak működési zavaraira.
- Depresszió: Antidepresszánsok, mint például a szerotonin-visszavétel gátlók (SSRI-k), fokozzák a szerotonin szintjét, ami cAMP emelkedéshez és PKA aktivációhoz vezet az agyban. Ez hozzájárulhat a neuronális plaszticitás növeléséhez és a hangulat javulásához.
- Parkinson-kór: A dopaminerg neuronok degenerációjával járó Parkinson-kórban a cAMP szint csökkenése is megfigyelhető, ami befolyásolja a neuronok túlélését és működését.
- Alzheimer-kór: Az amiloid-béta (Aβ) felhalmozódás, amely az Alzheimer-kór egyik jellemzője, gátolhatja a cAMP jelátvitelt, hozzájárulva a kognitív funkciók romlásához.
A PDE4 gátlók, amelyek emelik a cAMP szintet az agyban, potenciális kognitív enhancer és antidepresszáns hatásúak lehetnek, és jelenleg is folynak velük kapcsolatos klinikai vizsgálatok.
Fertőző betegségek és gyulladás
Számos bakteriális toxin közvetlenül célozza meg a cAMP jelátviteli rendszert, hogy befolyásolja a gazdaszervezet sejtjeinek működését. Például a kolera toxin (Vibrio cholerae) és a pertussis toxin (Bordetella pertussis) irreverzibilisen aktiválja a Gs fehérjét (kolera toxin) vagy gátolja a Gi fehérjét (pertussis toxin), ami tartósan magas cAMP szintet eredményez a bélhámsejtekben. Ez súlyos víz- és elektrolit-vesztéshez vezet, ami a kolera és a szamárköhögés fő tüneteiért felelős.
A gyulladásos betegségekben, mint az asztma, COPD, rheumatoid arthritis, a cAMP jelátvitel modulálása terápiás lehetőségeket kínál. A cAMP emelkedése gyulladáscsökkentő hatású, gátolja az immunsejtek aktiválódását és a gyulladásos mediátorok felszabadulását. A PDE4 gátlók, mint a roflumilaszt, már alkalmazásban vannak COPD kezelésében, kihasználva ezt a gyulladáscsökkentő hatást.
A cAMP jelátvitel diszregulációja nem csupán egy tünet, hanem gyakran a betegség alapvető mozgatórugója, amelynek megértése kulcsfontosságú a célzott terápiák kifejlesztésében.
Összességében a cAMP jelátviteli útvonalak patofiziológiai szerepének mélyebb megértése alapvető fontosságú a modern orvostudomány számára. A célzott gyógyszerfejlesztés, amely az adenilát-cikláz izoformákra, a specifikus PDE-kre vagy a cAMP effektorokra irányul, ígéretes utakat nyithat számos, jelenleg nehezen kezelhető betegség hatékonyabb terápiájához.
Farmakológiai moduláció: cAMP-vel kapcsolatos gyógyszerek

A cAMP jelátviteli rendszer alapvető szerepe a sejtfiziológiában és patofiziológiában logikusan vezette a kutatókat arra, hogy a rendszer komponenseit farmakológiai célpontként azonosítsák. A cAMP szintjének vagy effektorainak modulálása rendkívül hatékony terápiás stratégiákat kínál számos betegség kezelésében. A gyógyszerek fejlesztése ezen a területen a G-fehérje-kapcsolt receptorok agonistáira és antagonistáira, az adenilát-cikláz modulátoraira és különösen a foszfodiészteráz (PDE) gátlókra összpontosít.
G-fehérje-kapcsolt receptor (GPCR) agonisták és antagonisták
Mivel a cAMP termelést nagyrészt a GPCR-ek aktivációja szabályozza, a GPCR-eket célzó gyógyszerek közvetlenül befolyásolják a cAMP szintjét. A GPCR-ek a legnagyobb gyógyszercélpont családot alkotják, és számos, már forgalomban lévő gyógyszer hatásmechanizmusa a cAMP útvonalon keresztül érvényesül.
- Béta-adrenerg agonisták: Ezek a gyógyszerek, mint például a szalbutamol vagy a formoterol, a béta-adrenerg receptorokhoz kötődnek, aktiválják a Gs fehérjét és az adenilát-ciklázt, növelve a cAMP szintet. A hörgők simaizomsejtjeiben ez bronchodilatációt okoz, ezért asztma és COPD kezelésében alkalmazzák őket. A szívben a dobutamin, egy béta-1 agonista, pozitív inotróp hatású, és akut szívelégtelenségben használják.
- Béta-adrenerg antagonisták (béta-blokkolók): Ezek a gyógyszerek, mint a propranolol vagy a metoprolol, blokkolják a béta-adrenerg receptorokat, gátolva az adrenalin és noradrenalin hatását. Ez csökkenti a cAMP termelést a szívben, ami lassítja a szívverést, csökkenti a kontrakciós erőt, és mérsékli a vérnyomást. Hipertónia, angina, aritmiák és szívelégtelenség kezelésében széles körben alkalmazzák őket.
- Glukagon receptor agonisták/antagonisták: A glukagon receptor antagonisták potenciális gyógyszerek lehetnek a 2-es típusú cukorbetegségben, mivel csökkentik a glukagon által kiváltott cAMP szint emelkedést a májban, és ezáltal a glükóz termelést.
Adenilát-cikláz aktivátorok és gátlók
Bár a legtöbb gyógyszer közvetetten, a GPCR-eken keresztül modulálja az adenilát-cikláz aktivitását, vannak olyan vegyületek is, amelyek közvetlenül hatnak az enzimen.
- Forskolin: Egy természetes eredetű diterpén, amely közvetlenül aktiválja a membránkötött adenilát-cikláz izoformákat, és széles körben használják kutatási eszközként a cAMP útvonal tanulmányozására. Potenciális terápiás alkalmazásai is vannak, például glaukóma kezelésében (csökkenti a szembelnyomást).
- Citoszolos adenilát-cikláz (sAC) gátlók: Az sAC, amely a bikarbonátra és kalciumra érzékeny, potenciális célpontot jelent a spermium motilitás szabályozásában (fogamzásgátlás) vagy bizonyos vesebetegségek kezelésében.
Foszfodiészteráz (PDE) gátlók
A PDE-k gátlása rendkívül sikeres farmakológiai stratégia, mivel lehetővé teszi a cAMP (és cGMP) szintjének célzott emelését különböző szövetekben. A PDE családok sokfélesége lehetővé teszi a szelektív gyógyszerek fejlesztését, amelyek minimalizálják a mellékhatásokat.
- Nem szelektív PDE gátlók:
- Koffein és teofillin: Ezek a metilxantinok nem szelektíven gátolják számos PDE izoformát. A koffein központi idegrendszeri stimuláns hatása részben a cAMP szint emelésén keresztül valósul meg. A teofillint korábban asztma kezelésére használták bronchodilatátor és gyulladáscsökkentő hatása miatt, de szűk terápiás ablaka miatt ma már ritkábban alkalmazzák.
- Szelektív PDE gátlók:
- PDE3 gátlók (pl. milrinon, cilostazol): A milrinont akut szívelégtelenségben alkalmazzák a szívizom kontrakciójának fokozására. A cilostazolt perifériás artéria betegségben használják az erek tágítására és a vérlemezkék aggregációjának gátlására, mivel emeli a cAMP szintet az ér simaizomsejtekben és a vérlemezkékben.
- PDE4 gátlók (pl. roflumilaszt, apremilaszt): A roflumilaszt a COPD súlyos formáinak kezelésére szolgál, gyulladáscsökkentő hatása miatt. Az apremilaszt pikkelysömör és pszoriázisos ízületi gyulladás kezelésére használatos. Ezek a gyógyszerek a cAMP szint emelésén keresztül gátolják a gyulladásos citokinek termelését az immunsejtekben.
- PDE5 gátlók (pl. szildenafil, tadalafil): Bár elsősorban a cGMP jelátvitelt célozzák (a cGMP lebontását gátolva), fontos megjegyezni, hogy a cAMP és cGMP útvonalak gyakran keresztreagálnak. A PDE5 gátlókat erektilis diszfunkció és pulmonális hipertónia kezelésére használják.
- PDE10 gátlók: Jelenleg kutatás alatt állnak, mint potenciális gyógyszerek skizofrénia és Parkinson-kór kezelésére, mivel a PDE10A erősen expresszálódik az agy striátumában, és a dopaminerg jelátvitelt modulálja.
A cAMP jelátvitel farmakológiai modulációja egy dinamikusan fejlődő terület. A jövőbeli kutatások valószínűleg még specifikusabb PDE izoformákra, AKAP-okra, vagy akár az Epac fehérjékre ható gyógyszerek kifejlesztésére fókuszálnak majd. A cél az, hogy a cAMP jelátviteli rendszer finomhangolt modulálásával olyan terápiákat hozzanak létre, amelyek maximális hatékonyságot és minimális mellékhatásokat biztosítanak a betegek számára.







