

A sejtek, a biológiai élet alapegységei, rendkívül összetett és dinamikus rendszerek. A bennük zajló folyamatok, mint az anyagcsere, a szaporodás vagy az információfeldolgozás, mind precízen szabályozottak. Ezen szabályozás egyik legfundamentálisabb aspektusa a cellapotenciál, vagy más néven membránpotenciál. Ez a jelenség alapvetően meghatározza a sejtek működését, különösen az ideg- és izomsejtek esetében, de minden élő sejt rendelkezik vele. Egyszerűen fogalmazva, a cellapotenciál a sejt belseje és külseje közötti elektromos feszültségkülönbség, amelyet az ionok egyenlőtlen eloszlása és mozgása hoz létre a sejtmembránon keresztül.
Ahhoz, hogy megértsük a cellapotenciál lényegét, először is a sejt alapvető felépítésére és működésére kell tekintenünk. A sejt külső határa a sejtmembrán, amely egy vékony, féligáteresztő hártya. Ez a membrán nem csupán egy fizikai határ, hanem egy aktív, dinamikus felület, amely szabályozza, mi jut be a sejtbe és mi távozik onnan. A membránon keresztül történő anyagtranszport, különösen az ionok mozgása, kulcsszerepet játszik a potenciál kialakításában.
A sejtmembrán elsődleges alkotóeleme a foszfolipid kettős réteg. Ez a szerkezet hidrofób (víztaszító) belsővel és hidrofil (vízkedvelő) külsővel rendelkezik, ami hatékonyan elválasztja a sejt belső, vizes közegét (citoplazma) a külső, extracelluláris folyadéktól. A kettős réteg önmagában nagyrészt átjárhatatlan az ionok és a nagyobb, poláris molekulák számára. Azonban a membránba ágyazott fehérjék, mint az ioncsatornák és ionpumpák, lehetővé teszik a szelektív transzportot.
Az ionok szerepe és eloszlása
A cellapotenciál kialakulásához elengedhetetlen a különböző töltésű részecskék, az ionok egyenlőtlen eloszlása a sejtmembrán két oldalán. A legfontosabb ionok, amelyek hozzájárulnak ehhez a potenciálkülönbséghez, a következők:
- Nátrium-ionok (Na+): Jelentősen nagyobb koncentrációban vannak jelen a sejt külső oldalán.
- Kálium-ionok (K+): Jelentősen nagyobb koncentrációban találhatók a sejt belsejében.
- Klorid-ionok (Cl-): Magasabb koncentrációban vannak a sejt külsején.
- Kalcium-ionok (Ca2+): A sejt külső oldalán jóval koncentráltabbak.
Emellett a sejt belsejében számos nagy méretű, negatív töltésű molekula (pl. fehérjék, nukleinsavak) is található, amelyek nem tudnak átjutni a membránon, és hozzájárulnak a sejt belső negatív töltéséhez.
Ezek az ionkoncentráció-különbségek nem véletlenszerűek; aktív transzportmechanizmusok, például az Na+/K+-ATPáz pumpa tartja fenn őket. Ez a pumpa ATP energiáját felhasználva három Na+ iont pumpál ki a sejtből, és két K+ iont pumpál be a sejtbe minden ciklusban. Ez a mechanizmus nemcsak a koncentrációgradienseket tartja fenn, hanem közvetlenül is hozzájárul egy kis mértékű negatív töltés kialakulásához a sejt belsejében, mivel több pozitív töltést távolít el, mint amennyit bejuttat.
Elektrokémiai gradiens: a hajtóerő
Az ionok mozgását a sejtmembránon keresztül két fő erő hajtja: a koncentrációgradiens és az elektromos gradiens. Együtt alkotják az elektrokémiai gradienst.
- Koncentrációgradiens (kémiai gradiens): Az ionok igyekeznek a magasabb koncentrációjú helyről az alacsonyabb koncentrációjú helyre vándorolni, a diffúzió elve alapján. Például a K+ ionok, amelyek magasabb koncentrációban vannak a sejtben, hajlamosak kifelé áramlani.
- Elektromos gradiens: Az ionok az ellentétes töltésű területek felé vonzódnak, és a hasonló töltésű területektől taszítódnak. Például, ha a sejt belseje negatívabb, mint a külseje, a pozitív ionok (mint a K+ és Na+) hajlamosak befelé áramlani.
Ez a két erő egyensúlyozza egymást, és egy adott ion esetében az a pont, ahol a nettó áramlás megszűnik, az ion egyensúlyi potenciálja. Ezt az egyensúlyi potenciált az úgynevezett Nernst-egyenlet írja le, amely figyelembe veszi az ion koncentrációját a membrán két oldalán, valamint az ion töltését és a hőmérsékletet. Bár az egyenlet részletei meghaladják az „egyszerűen” magyarázat kereteit, fontos tudni, hogy minden egyes ionnak van egy saját egyensúlyi potenciálja, amelynél nincs nettó mozgása a membránon keresztül.
A cellapotenciál nem más, mint az ionok tánca a sejtmembránon keresztül, melyet a koncentráció és az elektromos erők bonyolult egyensúlya vezényel.
Nyugalmi membránpotenciál: a sejt alapszintje
A nyugalmi membránpotenciál az a stabil elektromos feszültségkülönbség, amely akkor mérhető a sejtmembrán két oldala között, amikor a sejt nem aktív, azaz nem kap ingert. Ez a potenciál általában negatív a sejt belsejében, és értéke fajtától és sejttípustól függően változik, jellemzően -40 mV és -90 mV között mozog. Az idegsejtek esetében például gyakran -70 mV körüli érték.
A nyugalmi potenciál kialakulásában kulcsszerepet játszik a sejtmembrán szelektív áteresztőképessége, különösen a K+ szivárgó csatornák (leak channels) jelenléte. Ezek a csatornák mindig nyitva vannak, és lehetővé teszik a K+ ionok szabad áramlását a membránon keresztül. Mivel a K+ koncentrációja a sejt belsejében sokkal magasabb, a K+ ionok a koncentrációgradiens hatására kifelé áramlanak a sejtből. Ez a kifelé áramló pozitív töltés a sejt belsejét egyre negatívabbá teszi, miközben a sejt külső oldalán pozitív töltést halmoz fel.
Ahogy a sejt belseje egyre negatívabbá válik, egy elektromos gradiens alakul ki, amely visszavonzza a pozitív K+ ionokat a sejtbe. Végül egy olyan pontra érkezünk, ahol a kémiai (koncentráció) gradiens, amely kifelé hajtja a K+ ionokat, egyensúlyba kerül az elektromos gradienssel, amely befelé vonzza őket. Ez az egyensúlyi pont nagyon közel van a K+ Nernst-potenciáljához, és ez határozza meg nagyrészt a nyugalmi membránpotenciált. Bár más ionok, mint a Na+ és Cl- is hozzájárulnak, a K+ áteresztőképessége a legdominánsabb a nyugalmi állapotban.
A Na+/K+-ATPáz pumpa létfontosságú a nyugalmi potenciál fenntartásában, nem pedig a közvetlen kialakításában. A pumpa folyamatosan dolgozik, hogy visszaállítsa az ionkoncentráció-különbségeket, amelyeket a szivárgó csatornákon keresztül történő ionmozgás folyamatosan igyekszik kiegyenlíteni. Ha a pumpa leállna, az iongradiensek lassan kiegyenlítődnének, és a nyugalmi potenciál megszűnne, ami a sejt működésképtelenségéhez vezetne.
Akciós potenciál: a sejtnyelv
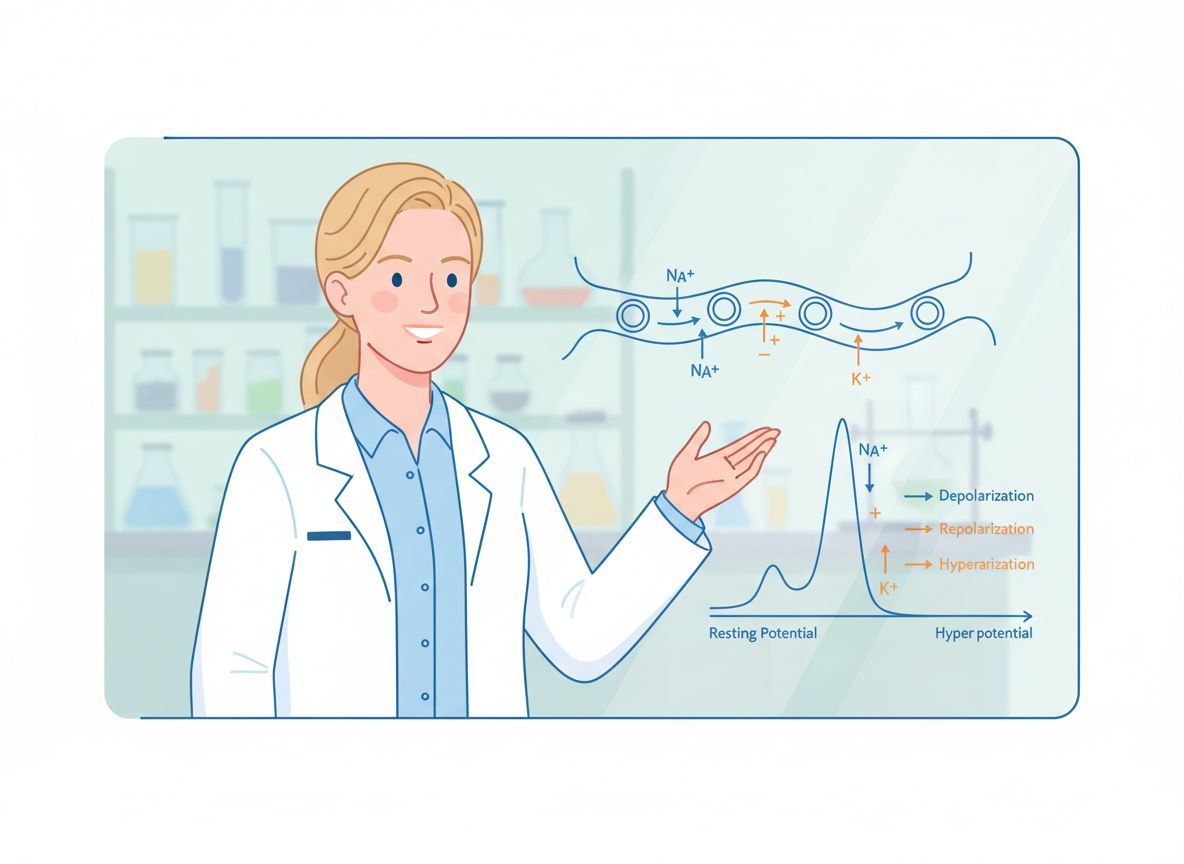
Míg a nyugalmi potenciál a sejt „alapállapota”, az akciós potenciál jelenti a sejt „kommunikációját” és „munkáját”. Az akciós potenciál egy gyors, tranziens, „mindent vagy semmit” típusú változás a membránpotenciálban, amely lehetővé teszi az információ gyors és távolsági továbbítását az idegsejtekben és az izomsejtek összehúzódását. Ez egy önszabályozó, hullámszerű jelenség, amely a sejtmembrán áteresztőképességének drámai és gyors változásain alapul.
Az akciós potenciál akkor jön létre, amikor egy sejt elegendő ingert kap ahhoz, hogy a membránpotenciálja elérjen egy bizonyos küszöbértéket. Ez a küszöbérték általában -50 és -55 mV között van. Amint a potenciál eléri ezt az értéket, egy sor feszültségfüggő ioncsatorna nyílik meg, ami elindítja az akciós potenciál fázisait.
Az akciós potenciál fázisai
- Depolarizáció (felszálló ág): Amint a membránpotenciál eléri a küszöböt, a feszültségfüggő Na+ csatornák gyorsan megnyílnak. Mivel a Na+ koncentrációja sokkal magasabb a sejt külsején, és a sejt belseje negatív, a Na+ ionok hatalmas sebességgel áramlanak be a sejtbe. Ez a pozitív töltés beáramlás rendkívül gyorsan pozitívvá teszi a membránpotenciált, akár +30-40 mV-ra is emelkedve. Ez a fázis felelős az akciós potenciál „csúcsáért”.
- Repolarizáció (leszálló ág): Röviddel a Na+ csatornák megnyílása után azok inaktiválódnak (bezáródnak, és nem nyithatók újra rövid ideig), és a feszültségfüggő K+ csatornák lassan megnyílnak. A K+ ionok, a magas belső koncentrációjuk miatt, kifelé áramlanak a sejtből. Ez a pozitív töltés kiáramlás visszaállítja a membránpotenciált a negatív tartományba.
- Hiperpolarizáció (utólagos hiperpolarizáció vagy túllövés): A K+ csatornák lassabban záródnak be, mint ahogy a Na+ csatornák inaktiválódtak. Ezért egy rövid ideig több K+ áramlik ki, mint amennyi a nyugalmi állapotban szivárogna, ami a membránpotenciált a nyugalmi potenciál alá viszi (pl. -80 mV-ra). Ezt nevezzük hiperpolarizációnak. Ezt követően a K+ csatornák bezáródnak, és a membránpotenciál visszatér a nyugalmi állapotba a K+ szivárgó csatornák és az Na+/K+-ATPáz pumpa működésének köszönhetően.
Az akciós potenciál „mindent vagy semmit” jellege azt jelenti, hogy ha az inger eléri a küszöböt, az akciós potenciál mindig azonos amplitúdóval és formával jön létre. Ha az inger nem éri el a küszöböt, nem jön létre akciós potenciál.
Refrakter periódusok
Az akciós potenciálok során két fontos refrakter periódus különböztethető meg:
- Abszolút refrakter periódus: Ez az az időszak, amikor a sejt semmilyen ingerre nem képes újabb akciós potenciált generálni. Ez a depolarizáció és a repolarizáció nagy részét fedi le, és a Na+ csatornák inaktivált állapotával magyarázható. Ez biztosítja, hogy az akciós potenciál csak egy irányba terjedjen, és hogy ne jöhessen létre túl gyorsan újabb akciós potenciál.
- Relatív refrakter periódus: Ebben az időszakban a sejt csak erősebb ingerre képes akciós potenciált generálni. Ez a hiperpolarizáció fázisát és a repolarizáció késői részét fedi le, amikor a K+ csatornák még nyitva vannak, és a membránpotenciál a nyugalmi potenciál alatt van. Ekkor több Na+ beáramlás szükséges a küszöb eléréséhez.
Az akciós potenciál terjedése
Az akciós potenciál nem marad azon a helyen, ahol keletkezett, hanem végigfut a sejtmembránon, az idegsejtek esetében az axonon. Ennek a terjedésnek a módja a neuron felépítésétől függ.
Vezetés nem mielinizált axonokon
A nem mielinizált axonokon az akciós potenciál folyamatosan terjed. Amikor egy területen akciós potenciál jön létre, a beáramló Na+ ionok depolarizálják a szomszédos mem területeket a küszöbértékig. Ez kiváltja az újabb feszültségfüggő Na+ csatornák megnyílását, és így az akciós potenciál hullámszerűen végigfut az axonon. Ez a folyamat viszonylag lassú, mivel a membrán minden pontján végbe kell mennie.
Vezetés mielinizált axonokon (szaltatórikus vezetés)
A legtöbb gerinces idegsejt axonja mielinhüvellyel van borítva. A mielin egy zsírban gazdag anyag, amelyet speciális sejtek (oligodendrociták a központi idegrendszerben, Schwann-sejtek a perifériás idegrendszerben) képeznek, és szigetelő réteget alkot az axon körül. A mielinhüvely megszakításokat mutat szabályos időközönként, ezeket nevezzük Ranvier-féle befűződéseknek.
A feszültségfüggő Na+ és K+ csatornák szinte kizárólag a Ranvier-féle befűződésekben koncentrálódnak. A mielin szigetelő hatása miatt az akciós potenciál „ugrásokkal” terjed egyik befűződésről a másikra. Amikor egy befűződésben akciós potenciál keletkezik, a beáramló Na+ ionok által keltett áram gyorsan végigfut a mielinizált szakaszon, és depolarizálja a következő befűződést a küszöbértékig. Ez a jelenség a szaltatórikus vezetés („ugró vezetés”), amely sokkal gyorsabbá teszi az ingerületvezetést, és energiahatékonyabb is, mivel kevesebb ionpumpa-munkára van szükség az iongradiensek helyreállításához.
Az akciós potenciál a sejt elektromos nyelve, amely lehetővé teszi a villámgyors kommunikációt az idegrendszerben és az izmok összehúzódását.
Szinaptikus átvitel: a sejtek közötti párbeszéd
Az idegsejtek nem önállóan működnek, hanem komplex hálózatokat alkotnak, amelyekben egymással kommunikálnak. Ez a kommunikáció a szinapszisokon keresztül történik, ahol az egyik neuron (preszinaptikus sejt) jelet küld egy másik neuronnak vagy egy effektor sejtnek (posztszinaptikus sejt).
Két fő típusú szinapszis létezik:
- Elektromos szinapszisok: Ezekben a szinapszisokban az ionok közvetlenül áramolhatnak az egyik sejtből a másikba, a réskapcsolatokon (gap junctions) keresztül. Ez nagyon gyors átvitelt biztosít, de kevésbé rugalmas.
- Kémiai szinapszisok: Ezek sokkal gyakoribbak, és a jelátvitel neurotranszmitterek (kémiai hírvivők) segítségével történik.
A kémiai szinapszisban az akciós potenciál eléri a preszinaptikus terminálist, ami a feszültségfüggő Ca2+ csatornák megnyílását váltja ki. A beáramló Ca2+ ionok hatására a neurotranszmittereket tartalmazó vezikulák fuzionálnak a preszinaptikus membránnal, és a neurotranszmitterek a szinaptikus résbe ürülnek. Ezek a molekulák a posztszinaptikus membránon lévő receptorokhoz kötődnek, ami ioncsatornák megnyílását vagy bezáródását okozza a posztszinaptikus sejtben. Ez a változás a posztszinaptikus membránpotenciálban posztszinaptikus potenciál (PSP) néven ismert.
A PSP-k két típusúak lehetnek:
- Excitációs posztszinaptikus potenciál (EPSP): Ezek depolarizálják a posztszinaptikus membránt, azaz közelebb viszik a küszöbértékhez, növelve az akciós potenciál kiváltásának valószínűségét. Általában Na+ vagy Ca2+ beáramlás okozza.
- Inhibíciós posztszinaptikus potenciál (IPSP): Ezek hiperpolarizálják a posztszinaptikus membránt, azaz távolabb viszik a küszöbértéktől, csökkentve az akciós potenciál kiváltásának valószínűségét. Általában Cl- beáramlás vagy K+ kiáramlás okozza.
A posztszinaptikus sejt integrálja az összes bejövő EPSP-t és IPSP-t (ezt nevezzük szummációnak, ami lehet térbeli vagy időbeli), és ha az eredő depolarizáció eléri a küszöböt, akkor újabb akciós potenciált generál. Ez a folyamat alapja az idegrendszer komplex információfeldolgozásának.
A cellapotenciál jelentősége a különböző sejttípusokban
Bár az akciós potenciál az ideg- és izomsejtekre jellemző, a membránpotenciál minden élő sejtben alapvető szerepet játszik, még ha nem is generálnak akciós potenciálokat.
| Sejttípus | A cellapotenciál szerepe | Példák |
|---|---|---|
| Idegsejtek (neuronok) | Információ továbbítása és feldolgozása az idegrendszerben. Az akciós potenciálok generálása és terjedése. | Szenzoros neuronok (érzékelés), motoros neuronok (mozgás), interneuronok (feldolgozás). |
| Izomsejtek (myocyták) | Izomösszehúzódás kiváltása. Az akciós potenciál eléri az izomsejtet, ami Ca2+ felszabadulást és kontrakciót eredményez. | Vázizom, szívizom, simaizom. |
| Mirigysejtek | Hormonok vagy más anyagok szekréciójának szabályozása. A membránpotenciál változásai kiválthatják a váladék felszabadulását. | Endokrin mirigyek (pl. hasnyálmirigy inzulin szekréciója), exokrin mirigyek. |
| Érzékelő (receptor) sejtek | Külső ingerek (fény, hang, nyomás, kémiai anyagok) elektromos jelekké alakítása (transzdukció). | Fotoreceptorok a szemben, szőrsejtek a fülben, ízlelőbimbók. |
| Szívizomsejtek | A szív ritmikus összehúzódásának generálása és terjesztése. Speciális pacemakerek generálják az akciós potenciálokat. | Szinuszcsomó, AV-csomó, Purkinje-rostok. |
| Növényi sejtek | Iontranszport, turgornyomás, válaszreakciók (pl. Mimosa pudica levélmozgása). | Gyökérsejtek, levélsejtek. |
A cellapotenciál és a betegségek

A membránpotenciál szabályozásának zavarai súlyos betegségekhez vezethetnek, amelyeket összefoglaló néven csatornopátiáknak nevezünk. Ezek az állapotok az ioncsatornák genetikai mutációiból vagy működési zavaraiból erednek, és érinthetik az idegrendszert, az izmokat, a szívet és más szerveket.
- Epilepszia: Az agyban fellépő rendellenes, túlzott és szinkronizált neuronális aktivitás, amelyet az ioncsatornák működésének zavarai okozhatnak, különösen a Na+ és K+ csatornáké.
- Szívritmuszavarok (arrhythmiák): A szívizomsejtek ioncsatornáinak rendellenességei okozhatják, ami a szív elektromos aktivitásának zavarához és szabálytalan szívveréshez vezet. Például a hosszú QT-szindróma a K+ csatornák hibájából adódik.
- Cisztás fibrózis: Egy genetikai betegség, amelyet a kloridcsatornák (CFTR – Cystic Fibrosis Transmembrane Conductance Regulator) hibás működése okoz. Ez befolyásolja a váladékok (nyálka, izzadság, emésztőnedvek) összetételét, ami súlyos szervi károsodásokhoz vezet.
- Izombénulás és izomgyengeség: Az izomsejtek ioncsatornáinak zavarai gátolhatják az izomösszehúzódást. Például a periodikus paralízis ritka formái a Na+ vagy K+ csatornák hibái miatt alakulnak ki.
- Migrén: Bizonyos migrénes formák, mint a familiáris hemiplegiás migrén, szintén ioncsatorna-mutációkkal hozhatók összefüggésbe.
- Sclerosis multiplex (SM): Bár nem közvetlenül ioncsatorna-betegség, az SM-ben a mielinhüvely pusztulása miatt az idegimpulzusok vezetése lelassul vagy teljesen leáll, ami súlyos neurológiai tüneteket okoz.
A toxinok és gyógyszerek is gyakran az ioncsatornákra hatnak, megváltoztatva a sejtpotenciált. Például a tetrodotoxin (TTX), a gömbhal méreganyaga, blokkolja a feszültségfüggő Na+ csatornákat, ezzel gátolva az akciós potenciálok kialakulását, ami bénuláshoz és halálhoz vezethet. Hasonlóképpen, a helyi érzéstelenítők is a Na+ csatornák gátlásával fejtik ki hatásukat, megakadályozva a fájdalomjelek továbbítását az idegekben.
A cellapotenciál mérése és vizsgálata
A cellapotenciál vizsgálata kulcsfontosságú a sejtfiziológia és a neurobiológia területén. Számos elektrofiziológiai technika létezik a membránpotenciál mérésére és manipulálására:
- Intracelluláris felvétel: Egy nagyon vékony üvegelektródát (mikroelektróda) vezetnek be a sejt belsejébe, és a sejt külső oldalán lévő referenciaelektródához képest mérik a feszültséget. Ez lehetővé teszi a nyugalmi potenciál és az akciós potenciálok közvetlen mérését.
- Patch-clamp technika: Ez a rendkívül kifinomult módszer lehetővé teszi egyetlen ioncsatorna áramának mérését. Egy üvegelektródát (patch-pipetta) szorosan ráfektetnek a sejtmembránra, és különböző konfigurációkban (pl. cell-attached, whole-cell, inside-out, outside-out) vizsgálhatják az ioncsatornák működését.
- Feszültség-clamp technika: Ezzel a módszerrel a membránpotenciált egy előre beállított értéken tartják, és mérik az áramot, amely ahhoz szükséges, hogy a potenciált ezen az értéken tartsák. Ez lehetővé teszi az ioncsatornák kinetikájának és feszültségfüggőségének részletes vizsgálatát.
- Áram-clamp technika: Ebben az esetben egy adott áramot injektálnak a sejtbe, és mérik az ebből eredő membránpotenciál-változásokat. Ezt használják az akciós potenciálok kiváltásának és terjedésének vizsgálatára.
Emellett léteznek nem invazív technikák is, mint az elektroenkefalográfia (EEG) az agyi aktivitás mérésére, az elektrokardiográfia (EKG) a szív elektromos működésének vizsgálatára, és az elektromiográfia (EMG) az izomaktivitás mérésére. Ezek a módszerek a populáció szintjén mérik a sok sejt együttes elektromos aktivitását, és fontos diagnosztikai eszközök.
Evolúciós perspektíva: miért alakult ki a cellapotenciál?
A cellapotenciál, különösen az akciós potenciál, az élet egyik leglenyűgözőbb evolúciós vívmánya. De miért volt szükség rá? A válasz a kommunikáció és az adaptáció szükségességében rejlik.
Az egysejtű szervezetek is fenntartanak membránpotenciált, ami alapvető fontosságú az anyagcseréhez, a tápanyagfelvételhez és a belső homeosztázis fenntartásához. Azonban a többsejtű szervezetek kialakulásával, különösen az idegrendszer és az izomrendszer megjelenésével, szükségessé vált a gyors és hatékony kommunikáció a távoli sejtek között. Képzeljük el egy ragadozó elől menekülő állatot: a látott képnek (fényingernek) gyorsan elektromos jellé kell alakulnia, továbbítódnia kell az agyba, ott feldolgozódnia, majd az agyból parancsnak kell indulnia az izmok felé, hogy azok összehúzódjanak és az állat elmeneküljön. Mindez pillanatok alatt zajlik, és ezt a sebességet csak az akciós potenciálok biztosíthatják.
A membránpotenciál lehetővé teszi a sejtek számára, hogy érzékeljék és válaszoljanak a környezeti változásokra. Az érzékelő sejtek a külső ingereket (fény, hang, hőmérséklet, kémiai anyagok) elektromos jelekké alakítják át, amelyek az idegrendszerbe továbbítódva információt szolgáltatnak a világról. Ez az ingerületátalakítás (transzdukció) alapja az érzékelésnek.
Az ioncsatornák és ionpumpák evolúciója, amelyek a cellapotenciált létrehozzák és fenntartják, lehetővé tette a komplex, koordinált viselkedés kialakulását. Az „elektromos” nyelven kommunikáló sejtek rendkívül gyorsan tudnak alkalmazkodni a változó körülményekhez, ami alapvető fontosságú a túléléshez és a fajok fejlődéséhez. A mielinhüvely megjelenése tovább gyorsította az ingerületvezetést, lehetővé téve még komplexebb idegrendszerek és gyorsabb reakciók kialakulását.
A cellapotenciál tehát nem csupán egy fizikai jelenség, hanem az élet egyik legősibb és leginkább alapvető mechanizmusa. A molekuláris szintű ionmozgásoktól a komplex viselkedésformákig, minden szinten befolyásolja az élő szervezetek működését. Egyszerűnek tűnő alapelvei ellenére a mögötte rejlő mechanizmusok rendkívül kifinomultak és precízek, biztosítva a sejtek számára a túléléshez és a prosperáláshoz szükséges eszközöket.
A modern orvostudomány és biológia folyamatosan mélyíti el ismereteinket a cellapotenciálról, és az új felfedezések reményt adnak a csatornopátiák és más neurológiai, kardiológiai betegségek hatékonyabb kezelésére. A jelenség megértése kulcsfontosságú ahhoz, hogy jobban megértsük önmagunkat és az élővilág lenyűgöző komplexitását.







