
A földi élet alapját képező energiatermelés és anyagcsere-folyamatok közül kiemelkedő helyet foglal el a fotoszintézis. Ez az éltető mechanizmus, amely a napfény energiáját felhasználva szén-dioxidból és vízből szerves anyagokat, elsősorban szénhidrátokat szintetizál, nem csupán a növények, algák és bizonyos baktériumok létfenntartásához elengedhetetlen, hanem közvetetten a bolygó teljes élővilágának energiaellátását is biztosítja. A fotoszintézis két fő fázisra osztható: a fényfüggő és a fényfüggetlen reakciókra. Míg az előbbi a napenergia kémiai energiává (ATP és NADPH) alakításáért felel, addig az utóbbi, amelyet leggyakrabban Calvin-Benson-Bassham (CBB) ciklusnak, vagy egyszerűen Calvin-ciklusnak nevezünk, a légköri szén-dioxid fixálásáért és szerves molekulákká történő átalakításáért felelős. Ez a cikk a CBB ciklus mélyebb megértésére invitálja az olvasót, feltárva annak lényegét, lépéseit, kulcsfontosságú elemeit és a fotoszintézisben betöltött pótolhatatlan szerepét.
A CBB ciklus nem csupán egy egyszerű reakciólánc; egy komplex, ciklikusan ismétlődő biokémiai útvonal, amely a kloroplasztiszok sztrómájában zajlik. Ez a folyamat a földi élet szempontjából kritikus fontosságú, hiszen a szén-dioxid megkötésével alapozza meg a biomassza termelését, és ezáltal az egész ökoszisztéma működését. A ciklus során a szervetlen szén szerves vegyületekké alakul, amelyek később építőkövekként szolgálnak a komplexebb molekulák, mint például a glükóz, keményítő, cellulóz vagy fehérjék szintéziséhez. Nélküle a növények nem tudnának növekedni, a légkör szén-dioxid szintje drámaian megemelkedne, és a tápláléklánc alapjai omlanának össze.
A CBB ciklus felfedezésének története
A fotoszintézis részleteinek megértése hosszú tudományos kutatás eredménye. Bár a folyamat létezését már a 18. században sejtették, a szén-dioxid fixálásának mechanizmusára vonatkozó pontos ismeretek csak a 20. század közepén alakultak ki. A kulcsfontosságú áttörést Melvin Calvin és munkatársai, Andrew Benson és James Bassham érték el a Kaliforniai Egyetemen (Berkeley) az 1940-es és 1950-es években. Ők a radioaktív izotópokkal jelölt szén-dioxid (14CO2) és a papírkromatográfia alkalmazásával térképezték fel a szén útját a fotoszintézis sötét reakcióiban.
A kutatócsoport algákat (Chlorella és Scenedesmus) használt, amelyeket rövid ideig 14CO2-nek tettek ki, majd gyorsan elpusztították és extrahálták a bennük található szerves vegyületeket. A radioaktív vegyületek azonosításával lépésről lépésre rekonstruálták a szén-dioxid beépülésének útját. Felfedezték, hogy az első stabil szerves termék egy három szénatomos vegyület, a 3-foszfoglicerát (3-PGA). Ez a felismerés volt az alapja a ma CBB ciklusnak nevezett folyamat felvázolásának. Melvin Calvin 1961-ben kémiai Nobel-díjat kapott ezért a munkájáért, elismerve a fotoszintézis szén-dioxid fixálási útvonalának feltárásában elért úttörő eredményeit.
A fotoszintézis két fő fázisa: fényfüggő és fényfüggetlen reakciók
Mielőtt mélyebben belemerülnénk a CBB ciklus részleteibe, érdemes röviden áttekinteni a fotoszintézis egészét. A folyamat, ahogy már említettük, két fő szakaszra bontható:
- Fényfüggő reakciók: Ezek a reakciók a kloroplasztiszok tilakoid membránjaiban zajlanak, és közvetlenül igénylik a napfény energiáját. A fényenergiát pigmentek, mint például a klorofill nyelik el, majd ez az energia elektronok mozgását indítja el. Ennek eredményeként vízmolekulák hasadnak (fotolízis), oxigén szabadul fel, és két kulcsfontosságú energiatároló molekula keletkezik: az ATP (adenozin-trifoszfát) és a NADPH (nikotinamid-adenin-dinukleotid-foszfát). Az ATP egy univerzális energiavaluta, míg a NADPH egy redukálóerővel rendelkező molekula, amely hidrogént és elektronokat szállít. Ezek a molekulák a fényfüggetlen reakciók, azaz a CBB ciklus működéséhez nélkülözhetetlenek.
- Fényfüggetlen reakciók (CBB ciklus): Ezek a reakciók a kloroplasztiszok sztrómájában mennek végbe, és bár közvetlenül nem igényelnek fényt, a fényfüggő reakciók termékeire (ATP és NADPH) támaszkodnak. Itt történik a légköri szén-dioxid megkötése és szerves anyagokká való átalakítása, a glükóz szintézisének előkészítése. Ez a cikkünk fő témája.
A két fázis szoros kölcsönhatásban áll egymással, együttesen alkotva a fotoszintézis egészét. Az ATP és NADPH a fényfüggő reakciók „termékei”, amelyek „üzemanyagként” szolgálnak a CBB ciklus számára, lehetővé téve a szén-dioxid redukcióját és a szénhidrátok felépítését. A CBB ciklus során felhasznált ATP és NADPH visszaalakul ADP-vé és NADP+-vá, amelyek aztán visszatérnek a tilakoid membránokba, hogy újra feltöltődjenek a fényenergia segítségével.
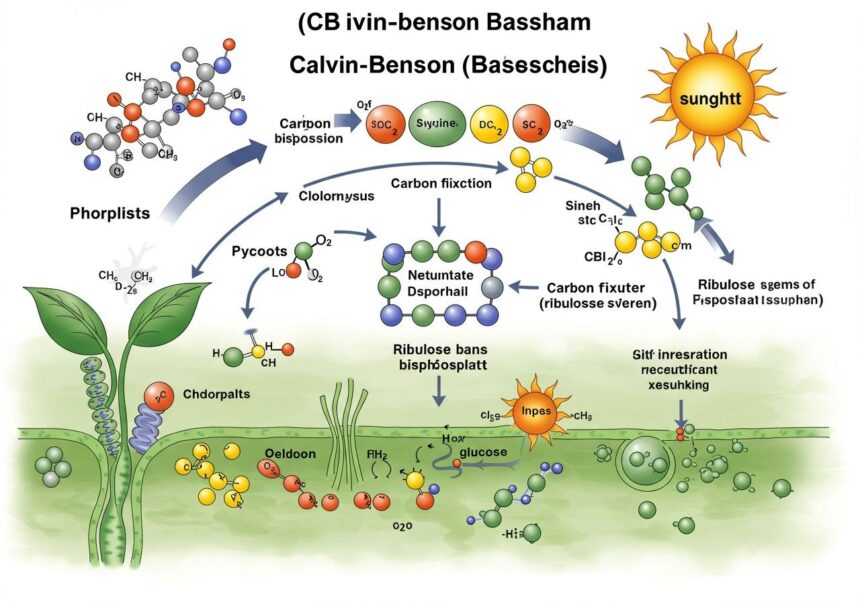
A CBB ciklus lépései: a szén útja a szerves anyagok felé
A CBB ciklus három fő szakaszra osztható, amelyek mindegyike alapvető fontosságú a szén-dioxid fixálása és a szénhidrátok szintézise szempontjából. Ezek a szakaszok ciklikusan ismétlődnek, biztosítva a folyamatos szén-dioxid megkötést.
1. Karboxilezés (szén-dioxid fixáció)
Ez a CBB ciklus első és talán legkritikusabb lépése, ahol a légköri szén-dioxid (CO2) beépül egy szerves molekulába. A folyamat kulcsszereplője a növényvilág legelterjedtebb és legfontosabb enzimje, a RuBisCO (ribulóz-1,5-biszfoszfát-karboxiláz/oxigenáz). Ez az enzim katalizálja a reakciót, amely során a CO2 egy öt szénatomos cukor-foszfát molekulához, a ribulóz-1,5-biszfoszfáthoz (RuBP) kapcsolódik.
A RuBisCO enzim karboxiláz aktivitása révén a CO2 kovalensen kötődik az RuBP-hez, egy instabil, hat szénatomos köztiterméket képezve. Ez a köztitermék azonnal két molekula 3-foszfoglicerát (3-PGA) formájában bomlik szét. A 3-PGA egy három szénatomos molekula, amely az első stabil szerves vegyület a CBB ciklusban, és amelynek felfedezése kulcsfontosságú volt a ciklus feltárásában. Minden egyes megkötött CO2 molekula két 3-PGA molekulát eredményez.
A RuBisCO enzim, bár létfontosságú, egyben a fotoszintézis egyik legkevésbé hatékony enzime is, mivel képes oxigént is kötni szén-dioxid helyett, ami a fotorespirációhoz vezet.
Ez a karboxilezési lépés alapozza meg a szerves anyagok szintézisét. A folyamat során a szervetlen szén atomok bekerülnek a biológiai rendszerekbe, ahol később energiatároló és építőanyagként szolgálnak. A RuBisCO tehát nem csupán egy enzim, hanem a földi élet egyik legfontosabb kapocsa a légkör és a biomassza között. A ciklus ezen szakaszában a szén-dioxid megkötésének hatékonysága számos tényezőtől függ, beleértve a CO2 koncentrációját, a hőmérsékletet és az enzim aktivitását.
2. Redukció (redukciós fázis)
A második szakaszban a 3-foszfoglicerát (3-PGA) molekulák redukálódnak, azaz energiát nyernek és átalakulnak egy magasabb energiatartalmú vegyületté, amely a szénhidrátok építőköveként szolgálhat. Ehhez a folyamathoz a fényfüggő reakciók során termelt energia és redukálóerő szükséges.
Először is, minden 3-PGA molekula egy ATP molekula felhasználásával foszforilálódik, azaz egy foszfátcsoportot kap, 1,3-biszfoszfoglicerátot (1,3-BPG) képezve. Ez a reakció az ATP hidrolízisével jár, amely során a nagy energiájú foszfátkötés energiája felszabadul és beépül a molekulába.
Ezt követően az 1,3-BPG redukálódik gliceraldehid-3-foszfáttá (G3P). Ez a redukció a NADPH molekulák által biztosított elektronok és protonok segítségével történik. A NADPH oxidálódik NADP+-ná, miközben az 1,3-BPG-t redukálja. A G3P egy három szénatomos cukor-foszfát, amely a CBB ciklus kulcsfontosságú terméke. Egy glükózmolekula szintéziséhez hat G3P molekula szükséges.
A G3P a CBB ciklus igazi „kimenete”. Ebből a molekulából épülnek fel a növények minden szerves anyagai, a cukroktól a fehérjékig és lipidekig.
A G3P molekulák sorsa többféle lehet:
- Egy részük (általában minden hat G3P-ből egy) kilép a ciklusból, és a kloroplasztiszban, vagy a citoplazmában más anyagcsere-folyamatokba lép. Ezekből a G3P molekulákból szintetizálódik a glükóz, a szacharóz, a keményítő és más komplex szénhidrátok. A glükóz a növények elsődleges energiaforrása, míg a keményítő hosszú távú energiatárolóként szolgál. A szacharóz pedig a cukor szállító formája a növényben.
- A G3P molekulák nagyobb része (általában öt a hatból) visszakerül a harmadik szakaszba, a regenerációs fázisba, hogy újra RuBP-vé alakuljon. Ez biztosítja a ciklus folyamatos működését.
3. Regeneráció (RuBP regeneráció)
A CBB ciklus harmadik és utolsó szakasza a ribulóz-1,5-biszfoszfát (RuBP) regenerálása. Ez a lépés elengedhetetlen ahhoz, hogy a ciklus folyamatosan működhessen, és újabb szén-dioxid molekulákat tudjon megkötni. A regeneráció során a korábban keletkezett gliceraldehid-3-foszfát (G3P) molekulákból újra RuBP képződik.
Ez a fázis egy komplex sorozatú reakciót foglal magában, amelyek során a három szénatomos G3P molekulákból öt szénatomos RuBP molekulák épülnek fel. Ehhez a folyamathoz további ATP energia felhasználása szükséges. Öt G3P molekulából, amelyek mindegyike három szénatomot tartalmaz (összesen 15 szénatom), három molekula RuBP képződik, amelyek mindegyike öt szénatomos (összesen 15 szénatom). Ez a szénatomok átrendeződését és foszforilációját magában foglaló folyamat biztosítja, hogy a szén-dioxid fixálásához szükséges kiindulási molekula mindig rendelkezésre álljon.
A regenerációs fázis során több enzim is közreműködik, amelyek a G3P molekulákat különböző cukor-foszfát köztitermékekké alakítják (pl. dihidroxiaceton-foszfát, fruktóz-6-foszfát, eritróz-4-foszfát, xilulóz-5-foszfát, szedoheptulóz-7-foszfát), míg végül kialakul a ribulóz-5-foszfát. Ezt a ribulóz-5-foszfátot aztán egy ATP molekula felhasználásával foszforilálja a foszforibulokináz enzim, így jön létre a ribulóz-1,5-biszfoszfát (RuBP). Ez a lépés zárja be a ciklust, és készíti fel a rendszert egy újabb szén-dioxid molekula megkötésére.
A regenerációs fázis energiaigényes, minden egyes RuBP molekula képzéséhez egy ATP molekula hidrolízisére van szükség. Ez ismételten rávilágít a fényfüggő reakciók által termelt ATP és NADPH elengedhetetlen szerepére a CBB ciklus fenntartásában.
Energiaigény és a fényfüggő reakciókkal való kapcsolat

A CBB ciklus nem egy spontán, energiafelszabadító folyamat; éppen ellenkezőleg, jelentős energiabefektetést igényel. Minden egyes megkötött CO2 molekula redukciójához és az RuBP regenerálásához a fényfüggő reakciók során termelt energiára van szükség. Pontosabban:
- 3 ATP molekula felhasználása történik. Ebből 2 ATP a 3-PGA foszforilálásához (1,3-BPG képzéséhez), 1 ATP pedig az RuBP regenerálásához szükséges.
- 2 NADPH molekula felhasználása történik, mindkettő a 1,3-BPG redukciójához (G3P képzéséhez).
Ez azt jelenti, hogy minden egyes szén-dioxid molekula beépítéséhez és a ciklus fenntartásához a növénynek jelentős mennyiségű ATP-t és NADPH-t kell termelnie a fényfüggő reakciók során. Ez a szoros kapcsolat a két fázis között biztosítja, hogy a fotoszintézis hatékonyan működjön, és a napenergia kémiai energiává alakulva végül szerves anyagok formájában raktározódjon.
A fényfüggő reakciók intenzitása közvetlenül befolyásolja a CBB ciklus sebességét. Erős fényben több ATP és NADPH termelődik, ami felgyorsítja a szén-dioxid fixálását. Gyenge fényben vagy sötétben az ATP és NADPH szintje lecsökken, ami lelassítja vagy leállítja a CBB ciklust, még akkor is, ha elegendő szén-dioxid áll rendelkezésre. Ez a mechanizmus biztosítja, hogy a növény ne pazarolja az erőforrásait, és csak akkor végezzen szén-dioxid fixálást, amikor elegendő energia áll rendelkezésre.
A RuBisCO enzim kettős természete: karboxiláz és oxigenáz aktivitás
Ahogy már említettük, a RuBisCO enzim kulcsszerepet játszik a CBB ciklusban. Azonban ez az enzim nem kizárólag a szén-dioxidot képes megkötni. Kettős természete van: képes karboxilázként és oxigenázként is működni.
- Karboxiláz aktivitás: Ez a kívánt funkció, amikor a RuBisCO a szén-dioxidot (CO2) köti meg az RuBP-hez, két molekula 3-PGA-t eredményezve. Ez a fotoszintézis alapja.
- Oxigenáz aktivitás: Ez a kevésbé kívánatos, de elkerülhetetlen mellékreakció, amikor a RuBisCO a oxigént (O2) köti meg az RuBP-hez. Ennek eredményeként egy molekula 3-PGA és egy molekula két szénatomos foszfoglikolát keletkezik.
A foszfoglikolát egy mérgező vegyület, amelyet a növénynek le kell bontania. Ez a folyamat a fotorespiráció néven ismert, és energiaveszteséggel jár a növény számára, mivel oxigént használ fel és CO2-t bocsát ki, miközben nem termel ATP-t vagy NADPH-t. A fotorespiráció különösen problémás magas hőmérsékleten és alacsony CO2/O2 arány mellett (pl. ha a sztómák zárva vannak a vízpazarlás elkerülése érdekében, és az O2 felhalmozódik a levélben, miközben a CO2 elfogy).
A RuBisCO oxigenáz aktivitása a fotoszintézis hatékonyságának egyik fő korlátja. Az evolúció során azonban egyes növények stratégiákat fejlesztettek ki ennek a problémának a minimalizálására, ami elvezet minket a C3, C4 és CAM fotoszintézis típusokhoz.
A CBB ciklust befolyásoló tényezők
A CBB ciklus sebessége és hatékonysága számos külső és belső tényezőtől függ. Ezeknek a tényezőknek a megértése kulcsfontosságú a növények növekedésének és terméshozamának optimalizálásához, különösen a mezőgazdaságban.
- Szén-dioxid koncentráció: A légköri CO2 koncentráció az egyik legfontosabb korlátozó tényező. Mivel a CO2 a CBB ciklus alapvető szubsztrátja, annak alacsony szintje korlátozza a karboxilezés sebességét, még akkor is, ha elegendő fény és víz áll rendelkezésre. A magasabb CO2 koncentráció általában növeli a fotoszintézis sebességét, különösen a C3 növényeknél, mivel csökkenti a RuBisCO oxigenáz aktivitását.
- Fényintenzitás: Bár a CBB ciklus fényfüggetlen, közvetetten függ a fénytől a fényfüggő reakciók révén termelt ATP és NADPH miatt. Alacsony fényintenzitás esetén kevesebb ATP és NADPH termelődik, ami lelassítja a CBB ciklust. Magas fényintenzitás esetén a ciklus sebessége nő, amíg más tényezők (pl. CO2) nem válnak korlátozóvá.
- Hőmérséklet: A hőmérséklet befolyásolja az enzimek aktivitását, így a CBB ciklusban részt vevő enzimeket, beleértve a RuBisCO-t is. Az optimális hőmérséklet tartományon kívül az enzimek denaturálódhatnak (túl magas hőmérsékleten) vagy aktivitásuk csökkenhet (túl alacsony hőmérsékleten). Magas hőmérsékleten ráadásul a RuBisCO oxigenáz aktivitása is megnő, ami fokozza a fotorespirációt.
- Vízellátás: A vízhiány közvetetten befolyásolja a CBB ciklust. A növények vízpazarlásának elkerülése érdekében a sztómáikat bezárják, ami megakadályozza a CO2 felvételét a légkörből. Ezáltal a levél belsejében a CO2 koncentrációja csökken, míg az O2 koncentrációja nő, elősegítve a fotorespirációt és gátolva a CBB ciklust.
- Táplálékanyagok: A fotoszintézishez és a CBB ciklus működéséhez számos táplálékanyagra van szükség. Például a nitrogén kulcsfontosságú a RuBisCO és más enzimek szintéziséhez, a foszfor az ATP és a foszforilált cukrok alkotóeleme, míg a magnézium a klorofill és az enzimek kofaktora.
Ezen tényezők komplex kölcsönhatása határozza meg a növény fotoszintetikus hatékonyságát és végső soron növekedését. A környezeti stressz, mint a szárazság, hőhullám vagy tápanyaghiány, jelentősen csökkentheti a CBB ciklus működését, ami hátrányosan befolyásolja a növények termelékenységét.
C3, C4 és CAM fotoszintézis: a CBB ciklus adaptációi
A CBB ciklus az alapvető mechanizmus a szén-dioxid fixálására, de az evolúció során a növények különböző stratégiákat fejlesztettek ki a környezeti kihívásokra (különösen a szárazságra és a magas hőmérsékletre) való reagálásként. Ezek a stratégiák a CBB ciklus előtti, kiegészítő szén-dioxid fixációs mechanizmusokban nyilvánulnak meg.
C3 növények
A legtöbb növény, beleértve a búza, rizs, szója és szinte az összes fa, C3 növény. Nevüket arról kapták, hogy a szén-dioxid fixálásának első stabil terméke egy három szénatomos vegyület, a 3-foszfoglicerát (3-PGA), amely közvetlenül a RuBisCO által katalizált reakcióban keletkezik. A C3 növényekben a CBB ciklus a levél mezofillum sejtjeiben zajlik. Ezek a növények jól prosperálnak mérsékelt éghajlaton, ahol a hőmérséklet nem túl magas, és a vízellátás megfelelő. Azonban meleg, száraz körülmények között a fotorespiráció jelentős energiát pazarol el náluk, csökkentve a fotoszintetikus hatékonyságot.
C4 növények
A C4 növények (pl. kukorica, cukornád, cirok) egy speciális mechanizmust fejlesztettek ki a RuBisCO oxigenáz aktivitásának minimalizálására és a fotoszintézis hatékonyságának növelésére meleg, száraz környezetben. Ezek a növények a szén-dioxidot először egy négy szénatomos vegyület formájában fixálják, innen ered a „C4” elnevezés. A folyamat két elkülönült sejttípusban zajlik:
- Mezofillum sejtekben: Itt a PEP-karboxiláz enzim köti meg a CO2-t a foszfoenolpiruvát (PEP) molekulához, oxálacetátot (egy négy szénatomos vegyületet) képezve. A PEP-karboxiláz nem érzékeny az oxigénre, így hatékonyan fixálja a CO2-t alacsony koncentrációk mellett is. Az oxálacetát ezután más négy szénatomos savakká (pl. malát, aszpartát) alakul.
- Nyalábburok sejtekben: A négy szénatomos savak a mezofillum sejtekből a nyalábburok sejtekbe transzportálódnak, ahol dekarboxileződnek, azaz CO2-t bocsátanak ki. Ez a CO2 koncentráltan szabadul fel a nyalábburok sejtekben, ahol a CBB ciklus zajlik. A magas helyi CO2 koncentráció elnyomja a RuBisCO oxigenáz aktivitását, és maximalizálja a karboxiláz aktivitását, ezáltal minimalizálva a fotorespirációt.
Ez a mechanizmus a C4 növényeket rendkívül hatékonnyá teszi meleg, napos környezetben, mivel kevesebb vizet veszítenek a sztómák zárása miatt, és mégis magas fotoszintetikus sebességet tartanak fenn.
CAM növények (Crassulacean Acid Metabolism)
A CAM növények (pl. kaktuszok, ananász, agave) a legszárazabb környezetekhez alkalmazkodtak. Ők egy időbeli elkülönítést alkalmaznak a szén-dioxid fixálásában:
- Éjszaka: A CAM növények éjszaka nyitják ki sztómáikat, amikor a hőmérséklet alacsonyabb, és a párolgás mértéke kisebb. Ekkor a PEP-karboxiláz enzim segítségével CO2-t kötnek meg, és négy szénatomos savak (malát) formájában tárolják a vakuólumaikban.
- Nappal: Napközben a sztómák bezárulnak, minimalizálva a vízpazarlást. A tárolt malát a vakuólumokból felszabadul, dekarboxileződik, és a felszabaduló CO2 a kloroplasztiszokban zajló CBB ciklusba kerül. A nappali magas CO2 koncentráció ismét elnyomja a RuBisCO oxigenáz aktivitását.
A CAM növények rendkívül víztakarékosak, de fotoszintetikus sebességük általában alacsonyabb, mint a C3 vagy C4 növényeké, mivel a CO2 fixálás és a CBB ciklus nem tud egyszerre, maximális sebességgel zajlani.
Az alábbi táblázat összefoglalja a főbb különbségeket a három fotoszintézis típus között a CBB ciklus kontextusában:
| Jellemző | C3 növények | C4 növények | CAM növények |
|---|---|---|---|
| CO2 fixálás helye | Mezofillum sejtek | Mezofillum és nyalábburok sejtek | Mezofillum sejtek |
| CO2 fixálás ideje | Nappal | Nappal | Éjszaka |
| Első CO2 fixáló enzim | RuBisCO | PEP-karboxiláz | PEP-karboxiláz |
| Első stabil termék | 3-PGA (3C) | Oxálacetát (4C) | Oxálacetát (4C) |
| CBB ciklus helye | Mezofillum sejtek | Nyalábburok sejtek | Mezofillum sejtek |
| Fotorespiráció | Magas (melegben) | Alacsony/minimális | Alacsony/minimális |
| Vízfelhasználás hatékonysága | Alacsony | Magas | Nagyon magas |
| Jellemző környezet | Mérsékelt, nedves | Meleg, napos, száraz | Száraz, sivatagi |
| Példák | Búza, rizs, szója, fák | Kukorica, cukornád, cirok | Kaktuszok, ananász, agave |
Mindhárom fotoszintézis típusban a CBB ciklus a szén-dioxid fixálásának végső állomása, ahol a szervetlen szén szerves vegyületekké alakul. A C4 és CAM mechanizmusok lényegében „CO2 pumpaként” működnek, amelyek koncentrálják a szén-dioxidot a CBB ciklus működésének helyén, optimalizálva a RuBisCO működését és csökkentve a fotorespirációt.
A CBB ciklus ökológiai és globális jelentősége

A CBB ciklus jelentősége messze túlmutat a növényi sejtek biokémiáján. Globális szinten ez a folyamat a földi élet alapja, és kulcsszerepet játszik a bolygó klímájának és ökoszisztémáinak szabályozásában.
- Primer produkció alapja: A CBB ciklus felelős a primer produkció túlnyomó részéért a Földön, ami a fotoszintetikus szervezetek által termelt szerves anyagok mennyiségét jelenti. Ez a szerves anyag a tápláléklánc alapját képezi, közvetlenül vagy közvetve minden heterotróf élőlény (állatok, gombák, baktériumok nagy része) táplálékforrása. Nélküle nem létezne biomassza, és az ökoszisztémák összeomlanának.
- Légköri szén-dioxid szabályozása: A CBB ciklus a légkörből vonja ki a szén-dioxidot, és beépíti a növényi biomasszába. Ez a folyamat kulcsfontosságú a globális szénciklusban, segítve a légköri CO2 koncentrációjának szabályozását. A fotoszintézis csökkenti az üvegházhatású gázok szintjét, így mérsékli a klímaváltozást. Az erdőirtás és a talajdegradáció viszont csökkenti a bolygó szén-dioxid megkötő kapacitását, hozzájárulva a légköri CO2 növekedéséhez.
- Oxigén termelés: Bár az oxigén a fényfüggő reakciók mellékterméke, a CBB ciklus elengedhetetlen a fényfüggő reakciók folyamatos működéséhez. Ha a CBB ciklus nem használná fel az ATP-t és NADPH-t, a fényfüggő reakciók leállnának, és az oxigéntermelés is megszűnne. Így a CBB ciklus közvetetten hozzájárul a légköri oxigénszint fenntartásához, ami az aerob légzéshez elengedhetetlen.
- Energiaforrás: A CBB ciklus során szintetizált szénhidrátok nemcsak a növények saját energiaforrásai, hanem az egész tápláléklánc energiaalapját is biztosítják. Az állatok a növények fogyasztásával jutnak energiához, majd az állatok lebontásával a mikroorganizmusok is hozzájutnak az energiához. Ez az energiaáramlás a CBB ciklussal kezdődik.
- Biogeokémiai ciklusok: A szénciklus mellett a CBB ciklus más biogeokémiai ciklusokkal is összefügg, például a vízciklussal (transzspiráció), a nitrogénciklussal (fehérjeszintézis) és a foszforciklussal (ATP és foszfátok).
A CBB ciklus tehát nem csupán egy biokémiai útvonal, hanem a földi élet motorja, amely nélkülözhetetlen a bolygó ökológiai egyensúlyához és az emberiség fennmaradásához.
Jövőbeli kutatások és alkalmazások
A CBB ciklus mélyreható megértése lehetőségeket teremt a mezőgazdaság és a biotechnológia számára. A kutatók világszerte azon dolgoznak, hogy a fotoszintézis, és ezen belül a CBB ciklus hatékonyságát növeljék, különösen a klímaváltozás és a növekvő globális népesség kihívásainak fényében.
- A RuBisCO optimalizálása: Mivel a RuBisCO a fotoszintézis sebességét korlátozó tényező lehet, különösen a C3 növényeknél, a kutatók megpróbálják genetikailag módosítani az enzimet, hogy növeljék a karboxiláz aktivitását és csökkentsék az oxigenáz aktivitását. Ez jelentősen növelhetné a terméshozamot.
- C4 fotoszintézis bevezetése C3 növényekbe: Egy ambitciózus cél a C4 fotoszintézis géneinek beültetése olyan fontos C3 növényekbe, mint a rizs, hogy növeljék azok hatékonyságát meleg, száraz körülmények között. Ez egy rendkívül komplex feladat, de hatalmas potenciállal rendelkezik az élelmiszerbiztonság javításában.
- Mesterséges fotoszintézis: A CBB ciklus inspirálja a mesterséges fotoszintézis kutatását is, amelynek célja a napfény energiájának felhasználása szén-dioxidból üzemanyagok vagy más hasznos vegyületek előállítására. Ez a technológia kulcsfontosságú lehet a jövő energiaellátásában és a szén-dioxid kibocsátás csökkentésében.
- Klímaváltozással szembeni ellenálló képesség növelése: A CBB ciklus mechanizmusainak megértése segíthet a növények nemesítésében vagy génmódosításában, hogy ellenállóbbak legyenek a szárazsággal, hőstresszel és magas CO2 koncentrációval szemben. Ez létfontosságú az élelmiszertermelés fenntarthatósága szempontjából.
A CBB ciklus tehát nem csupán egy tankönyvi fejezet, hanem egy élő, dinamikus folyamat, amelynek további kutatása és megértése alapvető fontosságú a bolygó jövője szempontjából. A biokémiai részletek aprólékos elemzése és a globális ökológiai összefüggések felismerése egyaránt rávilágít a fotoszintézis és ezen belül a CBB ciklus pótolhatatlan értékére.







