
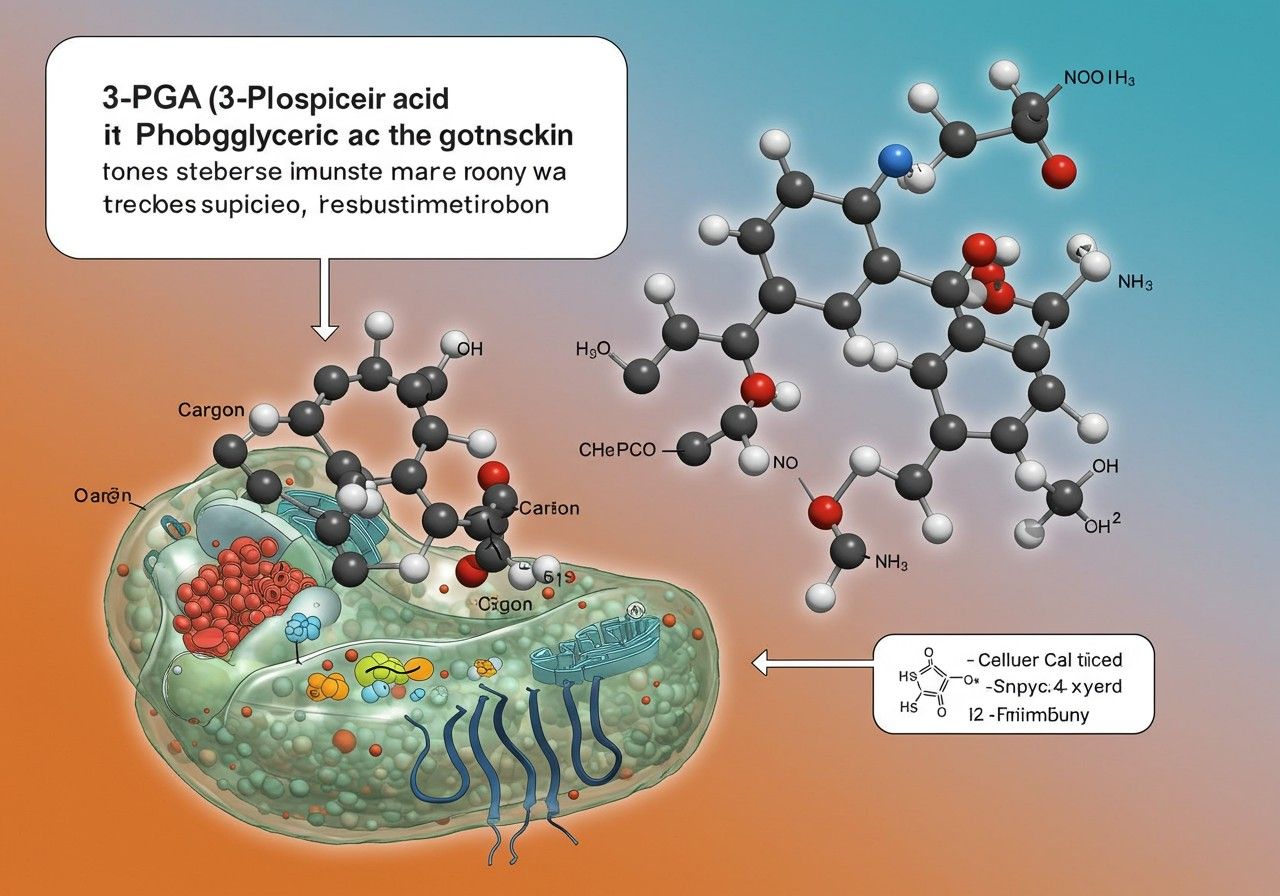
A 3-foszfoglicerát, vagy rövidebben 3-PGA, egy rendkívül fontos és sokoldalú molekula a sejtanyagcserében, amely számos alapvető biokémiai folyamat központi szereplője. Ez a relatíve egyszerű, három szénatomos vegyület a glikolízis, a fotoszintézis Calvin-ciklusa és a szerin bioszintézis kulcsfontosságú intermedierje, demonstrálva az élet bonyolult és összefüggő metabolikus hálózatát. Az élőlényekben betöltött fundamentális szerepe miatt a 3-PGA megértése elengedhetetlen a sejtbiológia és az anyagcsere folyamatainak mélyebb megismeréséhez.
A sejtanyagcsere komplex rendszere, amely magában foglalja a tápanyagok lebontását és az energia előállítását (katabolizmus), valamint az építőelemek szintézisét és a sejtek növekedését (anabolizmus), a 3-PGA révén szorosan összehangolt. Ez a molekula nem csupán egy egyszerű átmeneti termék, hanem egy olyan metabolikus csomópont, amely stratégiai ponton kapcsolja össze a szénhidrát-anyagcserét az aminosav-anyagcserével, és kulcsfontosságú a redukálóerő és az ATP termelésének szabályozásában.
A 3-PGA kémiai szerkezete és tulajdonságai
A 3-foszfoglicerát egy karbonsav, amelynek kémiai neve 2,3-dihidroxi-propionsav-3-foszfát. Szerkezetét tekintve egy három szénatomos glicerinsav molekula, amelynek harmadik (omega) szénatomjához egy foszfátcsoport kapcsolódik. A molekula egy karboxilcsoportot, két hidroxilcsoportot és egy foszfátcsoportot tartalmaz, amelyek mindegyike hozzájárul a vegyület biokémiai reaktivitásához és oldhatóságához. A foszfátcsoport jelenléte különösen fontos, mivel ez teszi lehetővé a molekula könnyű felismerését és átalakítását a különböző enzimek számára.
A 3-PGA egy poláris molekula, ami kiválóan oldódik vízben, így könnyen szállítódik a sejten belül a citoplazmában. A molekula foszfátcsoportja ionizálható, ami befolyásolja a molekula töltését a sejten belüli pH-viszonyoktól függően. Ez a tulajdonság kulcsfontosságú az enzimekkel való interakciók szempontjából, mivel az enzimek gyakran specifikusan ismernek fel töltött csoportokat a szubsztrátjaikon. A 3-PGA stabilitása is hozzájárul ahhoz, hogy hatékonyan részt vehessen a metabolikus útvonalakban, anélkül, hogy spontán módon lebomlana.
„A 3-PGA szerkezeti egyszerűsége és funkcionális sokoldalúsága teszi lehetővé, hogy a sejtanyagcsere egyik legfontosabb molekulájaként működjön.”
A foszfátcsoport nemcsak a molekula oldhatóságát és enzimatikus felismerését segíti, hanem a magas energiájú foszfátkötések kialakításában is szerepet játszhat a későbbi reakciók során. A 3-PGA molekula királis, ami azt jelenti, hogy létezik belőle egy tükörképi izomer is. Az élő rendszerekben jellemzően az L-izomer, az L-3-foszfoglicerát dominál, bár a D-izomer is előfordulhat bizonyos speciális metabolikus útvonalakon. Ez a sztereospecifikusság jellemző a biokémiai rendszerekre, ahol az enzimek rendkívül szelektívek a szubsztrátjaik térbeli elrendezésére.
A glikolízis kulcsfontosságú intermedierje
A glikolízis, azaz a cukorbontás az egyik legrégebbi és legáltalánosabb metabolikus útvonal a Földön, amely a glükóz lebontásával ATP-t és redukálóerőt termel. Ebben a tízlépéses folyamatban a 3-PGA egy kulcsfontosságú intermedier, amely a glikolízis második felében, az energia-termelő fázisban jelenik meg. A glikolízis során a glükóz két molekula piruváttá alakul át, miközben nettó két ATP és két NADH molekula keletkezik.
A 3-PGA a glikolízis hetedik lépésében keletkezik, amikor a 1,3-biszfoszfoglicerát molekula foszfátcsoportját a foszfoglicerát kináz enzim átviszi az ADP-re, ATP-t termelve. Ez a reakció az egyik első szubsztrát szintű foszforiláció a glikolízisben, ami közvetlenül ATP-t állít elő anélkül, hogy oxigénre vagy elektrontranszport láncra lenne szükség. Ez a mechanizmus rendkívül hatékony, különösen anaerob körülmények között, ahol az oxigénhiány miatt más ATP-termelő útvonalak korlátozottak.
A keletkezett 3-PGA ezután tovább alakul a glikolízisben. A nyolcadik lépésben a foszfoglicerát mutáz enzim izomerizálja, átalakítva 2-foszfogliceráttá. Ez a lépés egy intramolekuláris foszfátcsoport átvitelét jelenti, ahol a foszfátcsoport a harmadik szénatomról a másodikra kerül. Bár ez a reakció önmagában nem termel energiát, előkészíti a molekulát a következő, energia-termelő lépésre.
A 3-PGA tehát nemcsak egy egyszerű átmeneti termék, hanem egy olyan molekula, amelynek képződése és átalakulása szorosan szabályozott, és alapvető fontosságú a sejt energiaellátásának fenntartásában. A glikolízis ezen szakaszai demonstrálják, hogy a 3-PGA milyen stratégiai ponton helyezkedik el a sejt energetikai útvonalaiban, lehetővé téve a gyors és hatékony ATP-termelést a sejt szükségleteinek kielégítésére.
A Calvin-ciklus központi molekulája a fotoszintézisben
A fotoszintézis, a földi élet alapja, a napfény energiáját használja fel szerves anyagok előállítására szén-dioxidból és vízből. Ennek a folyamatnak a sötét szakasza, amelyet Calvin-ciklusnak nevezünk, a 3-PGA molekula körül forog. A Calvin-ciklus a kloroplasztiszok sztómájában zajlik, és a szén-dioxid fixálásáért felelős, azaz a CO2 beépítéséért a szerves molekulákba.
A Calvin-ciklus első és legfontosabb lépése a szén-dioxid fixálás. Ezt a reakciót a RuBisCO enzim (ribulóz-1,5-biszfoszfát-karboxiláz/oxigenáz) katalizálja, amely a ribulóz-1,5-biszfoszfát (RuBP) molekulához kapcsolja a CO2-t. Az így keletkező instabil hat szénatomos intermedier azonnal két molekula 3-foszfogliceráttá bomlik. Ez a reakció a fotoszintézis legkritikusabb lépése, mivel ezen keresztül kerül be a szén a szerves anyagok körforgásába.
A Calvin-ciklus következő fázisában a 3-PGA redukciója történik. Először a foszfoglicerát kináz ATP felhasználásával foszforilálja a 3-PGA-t, létrehozva az 1,3-biszfoszfoglicerátot. Ezt követően az NADH vagy NADPH által szolgáltatott redukálóerő segítségével (a növényekben jellemzően NADPH) a gliceraldehid-3-foszfát dehidrogenáz enzim redukálja az 1,3-biszfoszfoglicerátot gliceraldehid-3-foszfáttá (G3P). Ez a reakció kulcsfontosságú, mert ez az a pont, ahol a fényreakciók során termelt energia és redukálóerő beépül a szerves molekulák szintézisébe.
„A 3-PGA a fotoszintézis során a szén-dioxid fixálásának közvetlen terméke, alapja a növényi biomassza felépítésének.”
A keletkezett G3P molekulák egy része kilép a Calvin-ciklusból, hogy glükózt, szacharózt, keményítőt és más szerves vegyületeket szintetizáljon, amelyek a növény növekedéséhez és energiatárolásához szükségesek. A G3P másik része a Calvin-ciklus regenerációs fázisába lép, ahol bonyolult reakciósorozatokon keresztül újra ribulóz-1,5-biszfoszfáttá alakul, biztosítva a ciklus folyamatos működését. Így a 3-PGA nem csupán egy termék, hanem a fotoszintézis alapköve, amely nélkül a földi élet, ahogyan ismerjük, nem létezhetne.
A szerin bioszintézise és a 3-PGA kapcsolata

A szerin egy nem esszenciális aminosav, ami azt jelenti, hogy az emberi szervezet képes azt előállítani más metabolitokból. A szerin bioszintézise egy alapvető anabolikus útvonal, amely szorosan kapcsolódik a szénhidrát-anyagcseréhez, és a 3-PGA a kiindulási molekulája. Ez a kapcsolat kiemeli a 3-PGA metabolikus sokoldalúságát, hiszen nemcsak energia-termelő és szén-fixáló folyamatokban játszik szerepet, hanem az aminosav-szintézisben is.
A szerin bioszintézis három fő lépésben zajlik, amelyek mindegyike a 3-PGA-ból indul ki:
- Oxidáció: Az első lépésben a 3-PGA dehidrogenáz enzim oxidálja a 3-PGA-t, miközben NAD+ redukálódik NADH-vá. Ennek eredményeként 3-foszfopiruvát keletkezik. Ez a reakció egy alfa-ketosavat hoz létre, amely kulcsfontosságú az aminosav szintézisben.
- Transzamináció: A következő lépésben a foszfoszerin transzamináz enzim egy amincsoportot visz át egy glutamát molekuláról a 3-foszfopiruvátra. Ezzel a reakcióval foszfoszerin keletkezik, és alfa-ketoglutarát szabadul fel. Ez a transzaminációs reakció jellemző az aminosav-szintézisre, ahol a nitrogén beépítése történik.
- Defoszforiláció: Végül a foszfoszerin foszfatáz enzim hidrolizálja a foszfoszerint, eltávolítva a foszfátcsoportot, és tiszta szerint eredményezve. Ez a defoszforilációs lépés felszabadítja a kész aminosavat, amely aztán felhasználható fehérjeszintézisre vagy más metabolikus útvonalakra.
A szerin nem csupán a fehérjék építőköve, hanem prekurzora más fontos molekuláknak is, mint például a ciszteinnek, a glicinnek, a szfingolipideknek és a folátnak. A szerin bioszintézisének szabályozása tehát létfontosságú a sejt számára, és mivel a 3-PGA a kiinduló anyaga, a 3-PGA szintje és hozzáférhetősége közvetlenül befolyásolja ezt az útvonalat. Ez a metabolikus híd rávilágít arra, hogy a szénhidrát-anyagcsere és az aminosav-anyagcsere mennyire szorosan integrált a sejtben.
A 3-PGA és a glukoneogenezis
Míg a glikolízis a glükóz lebontásáról szól, addig a glukoneogenezis a glükóz szintézisét jelenti nem szénhidrát prekurzorokból, mint például laktátból, aminosavakból vagy glicerinből. Ez az útvonal különösen fontos az éhezés vagy hosszan tartó fizikai aktivitás idején, amikor a glikogénraktárak kimerülnek, és a szervezetnek folyamatosan glükózt kell biztosítania az agynak és a vörösvértesteknek. A 3-PGA a glukoneogenezisben is kulcsfontosságú intermedier, tükrözve a glikolízisben betöltött szerepét, de fordított irányban.
A glukoneogenezis során a piruvátból kiindulva egy sor enzimreakció révén szintetizálódik a glükóz. A glikolízis számos lépése reverzibilis, de van néhány irreverzibilis lépés, amelyet a glukoneogenezisnek meg kell kerülnie. A 3-PGA a glikolízishez hasonlóan a középső szakaszban jelenik meg, de itt a gliceraldehid-3-foszfátból (G3P) alakul ki. A G3P, amely a fruktóz-1,6-biszfoszfát hasításából keletkezik, a glukoneogenezisben a trióz-foszfát izomeráz enzim segítségével dihidroxiaceton-foszfáttá (DHAP) izomerizálódik.
Ezt követően a G3P és DHAP kondenzálódik, fruktóz-1,6-biszfoszfátot képezve, majd a fruktóz-6-foszfát és végül a glükóz-6-foszfát keletkezik. A 3-PGA a glukoneogenezisben a G3P-ből indul ki, amely a glikolízishez képest fordított irányban alakul át. A gliceraldehid-3-foszfát dehidrogenáz és a foszfoglicerát kináz enzimek, amelyek a glikolízisben ATP-t termelnek, a glukoneogenezisben fordított irányban működnek, ATP és NADH felhasználásával alakítják át a 3-PGA-t G3P-vé.
Ez a reverzibilis szakasz teszi lehetővé, hogy a sejt gyorsan alkalmazkodjon a változó energiaigényekhez, és szükség esetén glükózt szintetizáljon. A 3-PGA tehát egy központi molekula a glükóz homeosztázisában, amely lehetővé teszi a szervezet számára, hogy fenntartsa a vércukorszintet még táplálékhiányos állapotban is. Ez a kettős szerep – a glikolízisben és a glukoneogenezisben – rávilágít a metabolikus útvonalak szoros összekapcsolódására és a 3-PGA központi jelentőségére a sejt anyagcseréjének szabályozásában.
A pentóz-foszfát út és a 3-PGA közvetett kapcsolatai
A pentóz-foszfát út (PPP), más néven hexóz-monofoszfát sönt, egy alternatív glükóz-anyagcsere útvonal, amelynek fő célja a NADPH termelése és a pentózok, különösen a ribóz-5-foszfát előállítása. A NADPH kulcsfontosságú a reduktív bioszintézis folyamatokhoz (pl. zsírsav- és szteroidszintézis) és az oxidatív stressz elleni védelemhez. A ribóz-5-foszfát pedig a nukleotidok és nukleinsavak (DNS, RNS) szintézisének alapanyaga. Bár a 3-PGA nem közvetlen intermedierje a PPP-nek, közvetett kapcsolatokon keresztül szorosan integrált a PPP és más metabolikus útvonalak között.
A pentóz-foszfát út két fő szakaszra osztható: az oxidatív és a nem-oxidatív szakaszra. Az oxidatív szakaszban a glükóz-6-foszfátból NADPH keletkezik. A nem-oxidatív szakaszban pedig a pentózok interkonverziója zajlik, amelyek glikolízis intermedierjeivé alakulhatnak vissza. A fruktóz-6-foszfát és a gliceraldehid-3-foszfát (G3P), amelyek a PPP nem-oxidatív szakaszának termékei, a glikolízisben is részt vesznek. Mivel a 3-PGA közvetlenül átalakítható G3P-vé a glikolízisben (és fordítva a glukoneogenezisben), ez a kapcsolat biztosítja a metabolikus rugalmasságot.
„A pentóz-foszfát út és a glikolízis közötti átjárás révén a 3-PGA közvetve hozzájárul a NADPH és a nukleotidok szintéziséhez, kulcsfontosságú a sejt védelmében és növekedésében.”
Ez az integráció lehetővé teszi a sejt számára, hogy a metabolikus igényeknek megfelelően irányítsa a szénáramlást. Ha a sejtnek több NADPH-ra van szüksége (pl. magas oxidatív stressz esetén), a glükóz-6-foszfát a PPP felé terelődik. Ha nukleotidokra van szükség, a PPP ribóz-5-foszfátot termel. Ha pedig energia (ATP) szükséges, a G3P és a fruktóz-6-foszfát visszatér a glikolízisbe, ahol a 3-PGA is szerepet játszik. Ez a hálózatos szerkezet biztosítja, hogy a sejt mindig optimálisan reagáljon a környezeti változásokra és a belső igényekre.
A 3-PGA tehát, bár nem közvetlen szereplője a PPP-nek, stratégiai elhelyezkedése a glikolízisben lehetővé teszi, hogy befolyásolja a PPP-vel kapcsolatos metabolikus folyamatokat. A glikolízis és a PPP közötti szoros kapcsolat révén a 3-PGA giroszkópként funkcionál, segítve a sejt anyagcseréjének egyensúlyban tartását a különböző metabolikus igények kielégítése érdekében.
A 3-PGA regulációja és enzimatikus átalakulásai
A 3-PGA metabolizmusa szigorúan szabályozott, hogy a sejt energia- és építőanyag-szükségleteit optimálisan kielégítse. Ennek a szabályozásnak a középpontjában azok az enzimek állnak, amelyek a 3-PGA szintézisét és átalakulását katalizálják. A kulcsfontosságú enzimek közé tartozik a foszfoglicerát kináz, a foszfoglicerát mutáz és a 3-PGA dehidrogenáz, amelyek mindegyike specifikus szerepet játszik a 3-PGA különböző metabolikus útvonalakba való beillesztésében.
A foszfoglicerát kináz (PGK) a glikolízis és a glukoneogenezis reverzibilis enzime. A glikolízisben ATP-t termel a 1,3-biszfoszfoglicerátból 3-PGA-t képezve, míg a glukoneogenezisben ATP-t fogyaszt a 3-PGA foszforilálásához. Az enzim aktivitását az ATP/ADP arány és a szubsztrátok (1,3-biszfoszfoglicerát és ADP) koncentrációja befolyásolja. Magas ATP szint gátolhatja az enzimet a glikolízis irányában, míg alacsony ATP szint aktiválja. Ez a mechanizmus biztosítja, hogy a sejt energiaállapotától függően a glikolízis vagy a glukoneogenezis legyen domináns.
A foszfoglicerát mutáz (PGM) a 3-PGA és a 2-foszfoglicerát közötti izomerizációt katalizálja. Ez az enzim esszenciális a glikolízis és glukoneogenezis fenntartásához, mivel előkészíti a molekulát a következő, magas energiájú foszfátkötést tartalmazó intermedier (foszfoenolpiruvát) képződéséhez. A PGM aktivitását a 2,3-biszfoszfoglicerát (2,3-BPG) szintje is befolyásolja, amely egy alloszterikus aktivátor. A 2,3-BPG szintje a vörösvértestekben különösen fontos, mivel szabályozza a hemoglobin oxigénkötő képességét.
A 3-PGA dehidrogenáz (PHGDH) a szerin bioszintézis első és elkötelezett lépését katalizálja, a 3-PGA oxidációját 3-foszfopiruváttá. Ez az enzim kulcsfontosságú a szerin és a belőle származó más molekulák (pl. glicin, cisztein) termelésében. A PHGDH aktivitását alloszterikusan gátolja a szerin, ami egy klasszikus végtermék-gátlási mechanizmus. Ez biztosítja, hogy a sejt ne termeljen feleslegesen szerint, ha már elegendő mennyiség áll rendelkezésre.
Ezen enzimek mellett más faktorok, mint például a hormonális szabályozás és a génexpresszió is befolyásolják a 3-PGA metabolizmusát. Például az inzulin és a glukagon hormonok a glikolízis és glukoneogenezis kulcsenzimeit szabályozzák, közvetve hatva a 3-PGA szintjére is. A génexpresszió szintjén a sejtek képesek növelni vagy csökkenteni az adott enzimek mennyiségét a metabolikus igényeknek megfelelően, biztosítva a hosszú távú adaptációt.
Ez a komplex szabályozási hálózat garantálja, hogy a 3-PGA szintje és metabolikus áramlása mindig optimális legyen a sejt aktuális állapotához és környezetéhez képest, fenntartva a metabolikus homeosztázist.
A 3-PGA mint jelzőmolekula és stresszválasz

A 3-PGA nem csupán egy metabolikus intermedier, hanem egyre inkább felismerik, hogy jelzőmolekulaként is funkcionálhat, amely befolyásolja a sejtek stresszre adott válaszát és a génexpressziót. Ez a jelzőszerep rávilágít a metabolitok és a sejtes jelátviteli útvonalak közötti szoros kapcsolatra, és arra, hogy a metabolikus állapot hogyan kommunikálódik a sejtmaggal és a génexpressziós gépezettel.
A 3-PGA szintjének változása a sejten belül metabolikus stresszre utalhat. Például, ha a glikolízis felgyorsul, vagy a szerin bioszintézise megnövekszik, a 3-PGA szintje is emelkedhet. Ez az emelkedés aktiválhat bizonyos jelátviteli útvonalakat, amelyek a sejt adaptív válaszait koordinálják. Kutatások kimutatták, hogy a 3-PGA befolyásolhatja a redox egyensúlyt és az oxidatív stresszre adott válaszokat, mivel a szerin bioszintézise révén hozzájárul a glutation szintéziséhez, amely egy fontos antioxidáns.
„A 3-PGA nem csupán egy építőelem, hanem egy kommunikátor is, amely a sejt metabolikus állapotát jelzi, befolyásolva a stresszválaszt és a génexpressziót.”
Emellett a 3-PGA hatással lehet a transzkripciós faktorok aktivitására is. Egyes tanulmányok szerint a 3-PGA képes modulálni az mTOR (mammalian Target of Rapamycin) útvonalat, amely kulcsfontosságú a sejt növekedésének, proliferációjának és anyagcseréjének szabályozásában. Az mTOR aktivitása szorosan kapcsolódik a tápanyagellátottsághoz, és a 3-PGA szintje jelezheti a szénhidrát- és aminosav-elérhetőséget, így befolyásolva az mTOR jelátvitelt.
A növényekben a 3-PGA szintje szintén kulcsfontosságú a stresszválaszban. A fotoszintézis során a CO2 fixálásának első stabil termékeként a 3-PGA szintje közvetlenül tükrözi a fotoszintetikus aktivitást és a szén-dioxid elérhetőségét. Aszály, magas hőmérséklet vagy egyéb környezeti stressz esetén, amikor a fotoszintézis hatékonysága csökken, a 3-PGA termelése is változhat, ami jelzéseket indíthat el a növényben a stresszre való adaptációra.
Ezek a felfedezések új dimenziót nyitnak a metabolitok biológiai szerepének megértésében, és rávilágítanak arra, hogy a 3-PGA nemcsak egy passzív résztvevője a metabolikus útvonalaknak, hanem aktív szerepet játszik a sejtek környezeti kihívásokra adott válaszainak koordinálásában.
A 3-PGA szerepe a ráksejt-anyagcserében
A ráksejtek anyagcseréje jelentősen eltér a normális sejtekétől, gyakran mutatva egy jellegzetes metabolikus átprogramozást, amelyet Warburg-hatásnak nevezünk. Ez azt jelenti, hogy a ráksejtek még oxigén jelenlétében is preferenciálisan a glikolízist használják energiaforrásként, majd a keletkező piruvátot laktáttá fermentálják, ahelyett, hogy a mitokondriális oxidatív foszforilációt alkalmaznák. Ebben az átprogramozott anyagcserében a 3-PGA különösen fontos szerepet játszik.
A ráksejtek fokozott glikolízise megnöveli a 3-PGA termelését. Ez a megnövekedett 3-PGA szint nem csupán az ATP termelését segíti elő, hanem a bioszintetikus útvonalakhoz is biztosít prekurzorokat, amelyek elengedhetetlenek a gyorsan osztódó ráksejtek növekedéséhez és proliferációjához. A 3-PGA a szerin bioszintézis kiindulási anyaga, ahogy korábban említettük. A ráksejtek gyakran fokozott szerin szintézist mutatnak, ami létfontosságú a nukleotidok, a glutation és más fontos molekulák előállításához, amelyek támogatják a rákos növekedést és a stresszrezisztenciát.
A 3-PGA dehidrogenáz (PHGDH) enzim, amely a szerin bioszintézis első lépését katalizálja a 3-PGA-ból, gyakran túlműködik vagy amplifikálódik különböző ráktípusokban, például a melanomában és az emlőrákban. Ez a PHGDH túlexpresszió biztosítja a ráksejtek számára a bőséges szerin- és glicinellátást, amelyek elengedhetetlenek a gyors proliferációhoz, a DNS szintézishez és az antioxidáns védelemhez.
A 3-PGA metabolizmusának megértése és manipulálása ígéretes terápiás célpont lehet a rákellenes stratégiákban. A PHGDH gátlása például csökkentheti a szerin elérhetőségét a ráksejtek számára, ezáltal gátolva azok növekedését. A kutatók aktívan vizsgálják a 3-PGA metabolizmusát érintő molekuláris mechanizmusokat és a célzott terápiás beavatkozások lehetőségét, amelyek megzavarhatják a ráksejtek anyagcseréjét anélkül, hogy súlyosan károsítanák a normális sejteket.
A 3-PGA tehát nemcsak egy egyszerű metabolit, hanem egy kulcsfontosságú molekula, amelynek szerepe a ráksejt-anyagcserében alapvető fontosságú a tumorok növekedéséhez és túléléséhez. Ennek a szerepnek a mélyebb megértése új utakat nyithat a rákterápia fejlesztésében.
Evolúciós perspektívák: a 3-PGA ősi szerepe
A 3-PGA központi szerepe a modern sejtek anyagcseréjében arra utal, hogy ez a molekula már az élet korai szakaszában is alapvető jelentőséggel bírt. Az evolúciós biokémikusok úgy vélik, hogy a 3-PGA és a hozzá kapcsolódó metabolikus útvonalak, mint a glikolízis és a Calvin-ciklus, rendkívül ősi eredetűek, és már az első sejtekben is jelen voltak, vagy azok előfutáraiban.
A glikolízis, amely a 3-PGA-t is érinti, az egyik legősibb metabolikus útvonalnak számít. Mivel nem igényel oxigént, feltételezhető, hogy már az anoxikus, ősi Földön is működött, amikor az atmoszférában még nem volt szabad oxigén. A glikolízis szinte univerzális előfordulása minden élőlényben, a baktériumoktól az emberig, megerősíti ősi eredetét és alapvető fontosságát az energiaellátásban. A 3-PGA ebben az útvonalban kulcsfontosságú molekula az ATP termelésében.
Hasonlóképpen, a Calvin-ciklus, amelyben a 3-PGA a szén-dioxid fixálásának első stabil terméke, szintén egy ősi mechanizmus. Bár a fotoszintézis, ahogyan ma ismerjük, a cianobaktériumok megjelenésével vált dominánssá, az alapvető szén-fixáló mechanizmusok valószínűleg már korábban is léteztek, akár más redukáló ágenseket használva. A 3-PGA képződése a szén-dioxidból egy rendkívül hatékony módja a szén beépítésének a szerves anyagokba, ami alapvető volt az élet fejlődéséhez.
„A 3-PGA univerzális jelenléte a glikolízisben és a fotoszintézisben egyértelműen jelzi, hogy ez a molekula az élet kialakulásának és evolúciójának egyik alapköve.”
A 3-PGA szerepe a szerin bioszintézisében is ősi kapcsolatokra utal. Az aminosavak szintézise a glikolízis intermedierjeiből egy hatékony módja volt a komplexebb biomolekulák előállításának az ősi sejtekben. Ez a metabolikus híd lehetővé tette a sejt számára, hogy a szénhidrát-anyagcseréből származó szénvázakat felhasználja a fehérjék építőköveinek előállítására, optimalizálva a korlátozott erőforrások felhasználását.
Az a tény, hogy a 3-PGA ennyire mélyen beépült ennyi alapvető metabolikus útvonalba, azt sugallja, hogy ez a molekula egyfajta metabolikus „központ” volt már az élet korai szakaszában is. A molekula egyszerű szerkezete, reaktivitása és a foszfátcsoport jelenléte valószínűleg ideálissá tette az ősi, kevésbé fejlett enzimrendszerek számára is. A 3-PGA tanulmányozása tehát nemcsak a modern sejtbiológia megértéséhez járul hozzá, hanem betekintést enged az élet eredetébe és evolúciós történetébe is.
A 3-PGA analógjai és gyógyszerészeti vonatkozásai
A 3-PGA központi szerepe a sejtanyagcserében, különösen a ráksejtek metabolikus átprogramozásában, felkeltette a gyógyszerkutatók érdeklődését. A molekula analógjainak vagy az azt átalakító enzimek gátlóinak kifejlesztése ígéretes terápiás stratégiákat kínálhat különböző betegségek, különösen a rák kezelésében. A cél az, hogy szelektíven megzavarják a beteg sejtek metabolizmusát anélkül, hogy súlyosan károsítanák az egészséges sejteket.
Az egyik legfontosabb célpont a 3-PGA dehidrogenáz (PHGDH) enzim, amely a szerin bioszintézis első lépését katalizálja 3-PGA-ból. Mivel a PHGDH gyakran túlműködik bizonyos ráktípusokban, a PHGDH-t gátló molekulák fejlesztése aktív kutatási terület. Ezek a gátlók elméletileg csökkenthetnék a szerin és glicin elérhetőségét a ráksejtek számára, ezáltal gátolva azok növekedését és proliferációját. Már fejlesztés alatt állnak olyan kis molekulájú gátlók, amelyek specifikusan célozzák a PHGDH-t, és ígéretes eredményeket mutatnak preklinikai vizsgálatokban.
A PHGDH gátlása azonban kihívásokat is rejt magában. A szerin esszenciális aminosav az agy számára, és a szerin bioszintézisének teljes gátlása potenciálisan mellékhatásokhoz vezethet. Ezért a gyógyszerfejlesztés során a szelektivitás és a biztonságosság kulcsfontosságú. A kutatók olyan gátlókat keresnek, amelyek specifikusan a ráksejtekben hatnak, vagy amelyek dózisa optimalizálható a mellékhatások minimalizálása érdekében.
A 3-PGA metabolizmusának más aspektusai is potenciális terápiás célpontok lehetnek. Például a foszfoglicerát mutáz (PGM) vagy a foszfoglicerát kináz (PGK) enzimek gátlása is befolyásolhatja a ráksejtek anyagcseréjét. Ezeknek az enzimeknek a gátlása megzavarhatja a glikolízist és a glukoneogenezist, megfosztva a ráksejteket az energiaforrásuktól és a bioszintetikus prekurzoroktól.
| Célpont Enzim | Metabolikus Útvonal | Potenciális Terápiás Hatás |
|---|---|---|
| 3-PGA dehidrogenáz (PHGDH) | Szerin bioszintézis | Ráksejt proliferáció gátlása, antioxidáns védelem csökkentése |
| Foszfoglicerát kináz (PGK) | Glikolízis, Glukoneogenezis | ATP termelés és szintézis zavarása |
| Foszfoglicerát mutáz (PGM) | Glikolízis, Glukoneogenezis | Metabolikus áramlás megzavarása, energiaellátás korlátozása |
A 3-PGA analógjai, amelyek versenghetnek az enzimek kötőhelyeiért, szintén érdekesek lehetnek. Ezek a molekulák úgy viselkedhetnek, mint a 3-PGA, de nem alakulnak tovább a metabolikus útvonalon, ezáltal blokkolva a normális metabolikus áramlást. Az ilyen típusú gyógyszerek fejlesztése bonyolult, mivel figyelembe kell venni a molekula bejutását a sejtekbe, stabilitását és szelektivitását.
Összességében a 3-PGA és a hozzá kapcsolódó enzimek célzása egy ígéretes megközelítés a gyógyszerfejlesztésben, különösen a rákterápia területén. A metabolikus útvonalak mélyebb megértése lehetővé teszi a specifikusabb és hatékonyabb terápiák kidolgozását, amelyek kihasználják a ráksejtek egyedi metabolikus sebezhetőségét.
A 3-PGA kutatása és jövőbeli irányai

A 3-PGA, mint a sejtanyagcsere központi molekulája, folyamatosan a kutatások fókuszában áll. A modern biokémiai, molekuláris biológiai és metabolomikai technikák fejlődése lehetővé teszi, hogy egyre mélyebben megértsük a 3-PGA szerepét nemcsak az alapvető metabolikus útvonalakban, hanem a komplex sejtes folyamatokban, mint például a jelátvitel, a génexpresszió és a betegségek patogenezise.
A jövőbeli kutatások egyik fő iránya a 3-PGA mint jelzőmolekula szerepének további feltárása. Hogyan érzékeli a sejt a 3-PGA szintjének változását? Milyen jelátviteli útvonalakon keresztül kommunikálja ez a molekula a metabolikus állapotot a génexpressziós gépezettel? A metabolitok és a transzkripciós faktorok közötti közvetlen interakciók feltárása új betekintést nyújthat a metabolikus szabályozásba és a betegségek kialakulásába.
A ráksejt-anyagcsere továbbra is kiemelt fontosságú terület marad. A 3-PGA metabolizmusának finomhangolása, különösen a PHGDH enzim működésének részletesebb vizsgálata, új célpontokat azonosíthat a rákterápiában. A kombinált terápiák, amelyek a PHGDH gátlását más rákellenes szerekkel ötvözik, ígéretesek lehetnek. Emellett a metabolikus képalkotó technikák, mint például az NMR vagy a tömegspektrometria, segíthetnek a 3-PGA szintjének non-invazív monitorozásában a tumorokban, ami prognosztikai vagy terápiás válaszmarkerként szolgálhat.
„A 3-PGA-val kapcsolatos kutatások folyamatosan bővítik tudásunkat a sejtanyagcsere komplexitásáról és új utakat nyitnak a betegségek kezelésében.”
A növényi anyagcsere területén a 3-PGA kutatása segíthet a fotoszintézis hatékonyságának növelésében. A klímaváltozás és a globális élelmiszerellátás kihívásai miatt létfontosságú a növények szén-dioxid fixáló képességének optimalizálása. A RuBisCO enzim, amely a 3-PGA-t termeli, híresen ineffektív, mivel oxigenáz aktivitással is rendelkezik. A 3-PGA termelését optimalizáló vagy a fotoszintézis mellékreakcióit csökkentő genetikai módosítások jelentősen javíthatják a terméshozamot.
A mikrobiális metabolizmusban is jelentős szerepet játszik a 3-PGA. A mikroorganizmusok metabolikus útvonalainak manipulálása biotechnológiai célokra, például bioüzemanyagok, gyógyszerek vagy ipari vegyületek előállítására, egyre nagyobb hangsúlyt kap. A 3-PGA metabolizmusának finomhangolása javíthatja bizonyos termékek hozamát a fermentációs folyamatokban.
A rendszerbiológia és a metabolomika integrált megközelítései alapvető fontosságúak lesznek a 3-PGA teljes metabolikus hálózatba való beágyazottságának megértésében. Nagy áteresztőképességű adatok elemzésével modellezni lehet a metabolikus áramlásokat, előre jelezni a metabolit szintek változásait és azonosítani a kulcsfontosságú szabályozási pontokat. Ez a megközelítés nemcsak a 3-PGA-ra, hanem a teljes sejtanyagcserére vonatkozóan is mélyebb betekintést nyújt.
Összességében a 3-PGA kutatása továbbra is izgalmas és dinamikus terület marad, amely alapvető biológiai kérdéseket feszeget, és potenciálisan új terápiás és biotechnológiai alkalmazásokhoz vezethet. A molekula egyszerűsége ellenére a benne rejlő komplexitás és sokoldalúság még hosszú ideig inspirálni fogja a tudósokat.







