
Minden élő szervezetnek energiára van szüksége a létezéshez, a növekedéshez, a mozgáshoz és a belső folyamatok fenntartásához. Gondoljunk csak egy izom összehúzódására, egy idegimpulzus továbbítására vagy akár a fehérjék szintézisére – mindezekhez az életfolyamatokhoz üzemanyagra van szükség. Az emberi test, és szinte minden komplex élőlény sejtjei ezt az energiát elsősorban egy molekula, az adenozin-trifoszfát (ATP) formájában tárolják és hasznosítják. Az ATP nem véletlenül kapta az ‘energiapénz’ elnevezést, hiszen ez a molekula szolgál a sejtek univerzális energiaforrásaként.
De hogyan termelődik ez a létfontosságú ATP? A válasz a sejtek mélyén, apró, de annál jelentősebb organellumokban, a mitokondriumokban rejlik. Itt zajlik a sejtlégzés utolsó, legfontosabb szakasza, amelyet légzési láncnak vagy elektrontranszport láncnak nevezünk. Ez a komplex biokémiai folyamat felelős az aerob szervezetek ATP-termelésének legnagyobb részéért, hatékonyan alakítva át a táplálékból származó energiát hasznosítható formává.
A légzési lánc megértése kulcsfontosságú ahhoz, hogy felfogjuk az élet energetikai alapjait. Ez nem csupán egy biokémiai reakciósorozat, hanem egy rendkívül finoman hangolt gépezet, amelynek hibátlan működése nélkülözhetetlen a sejtek, szövetek és az egész szervezet egészségéhez. Cikkünk célja, hogy a légzési lánc bonyolult mechanizmusát a lehető legegyszerűbben, mégis szakmailag hitelesen magyarázza el, feltárva annak minden apró részletét és óriási jelentőségét.
A sejtlégzés áttekintése: hol helyezkedik el a légzési lánc?
Mielőtt mélyebben belemerülnénk a légzési lánc részleteibe, érdemes áttekinteni a teljes sejtlégzés folyamatát, amelynek ez az utolsó, de leginkább termelékeny szakasza. A sejtlégzés egy gyűjtőfogalom, amely magában foglalja azokat az anyagcsere-folyamatokat, amelyek során a sejtek lebontják a szerves molekulákat (elsősorban glükózt) energiatermelés céljából. Ez a folyamat több lépcsőben zajlik, és mindegyiknek megvan a maga specifikus szerepe.
Az első szakasz a glikolízis, amely a sejtplazmában (citoplazmában) történik. Itt a glükóz egy tízlépéses folyamat során két piruvát molekulává alakul. Ez a szakasz oxigén jelenlétében és hiányában is végbemegy, és nettó 2 ATP-t és 2 NADH molekulát termel. A NADH (nikotinamid-adenin-dinukleotid) egy elektronhordozó molekula, amely a későbbiekben kulcsszerepet játszik az energiatermelésben.
Amennyiben oxigén áll rendelkezésre, a piruvát molekulák belépnek a mitokondriumba, ahol a következő lépések zajlanak. Először a piruvát oxidációja történik, melynek során a piruvát acetil-CoA-vá alakul, miközben CO2 szabadul fel és további NADH keletkezik. Ez az acetil-CoA ezután belép a Krebs-ciklusba (más néven citrátkörbe vagy trikarbonsav-ciklusba).
A Krebs-ciklus a mitokondrium mátrixában zajló körfolyamat, amely során az acetil-CoA teljesen oxidálódik, CO2-t termelve. Bár a Krebs-ciklus közvetlenül csak kevés ATP-t (pontosabban GTP-t, ami könnyen átalakul ATP-vé) termel, fő jelentősége abban rejlik, hogy nagy mennyiségű elektronhordozó molekulát, nevezetesen NADH-t és FADH2-t (flavin-adenin-dinukleotidot) generál. Ezek a redukált koenzimek hordozzák azokat a nagy energiájú elektronokat, amelyek a légzési láncban kerülnek majd felhasználásra.
És itt jön a képbe a légzési lánc, a sejtlégzés utolsó, de messze legtermelékenyebb szakasza. Ez a folyamat a mitokondrium belső membránjában zajlik, és az NADH-ból és FADH2-ből származó elektronok energiáját használja fel ATP szintézisére. Az aerob sejtlégzés során termelt ATP mintegy 90%-a ebben a szakaszban keletkezik, ami rávilágít a légzési lánc központi szerepére az energiaellátásban.
A sejtlégzés egy gondosan koreografált tánc, ahol a glikolízis, a Krebs-ciklus és a légzési lánc lépései harmonikusan illeszkednek egymáshoz, biztosítva a sejt folyamatos energiaellátását.
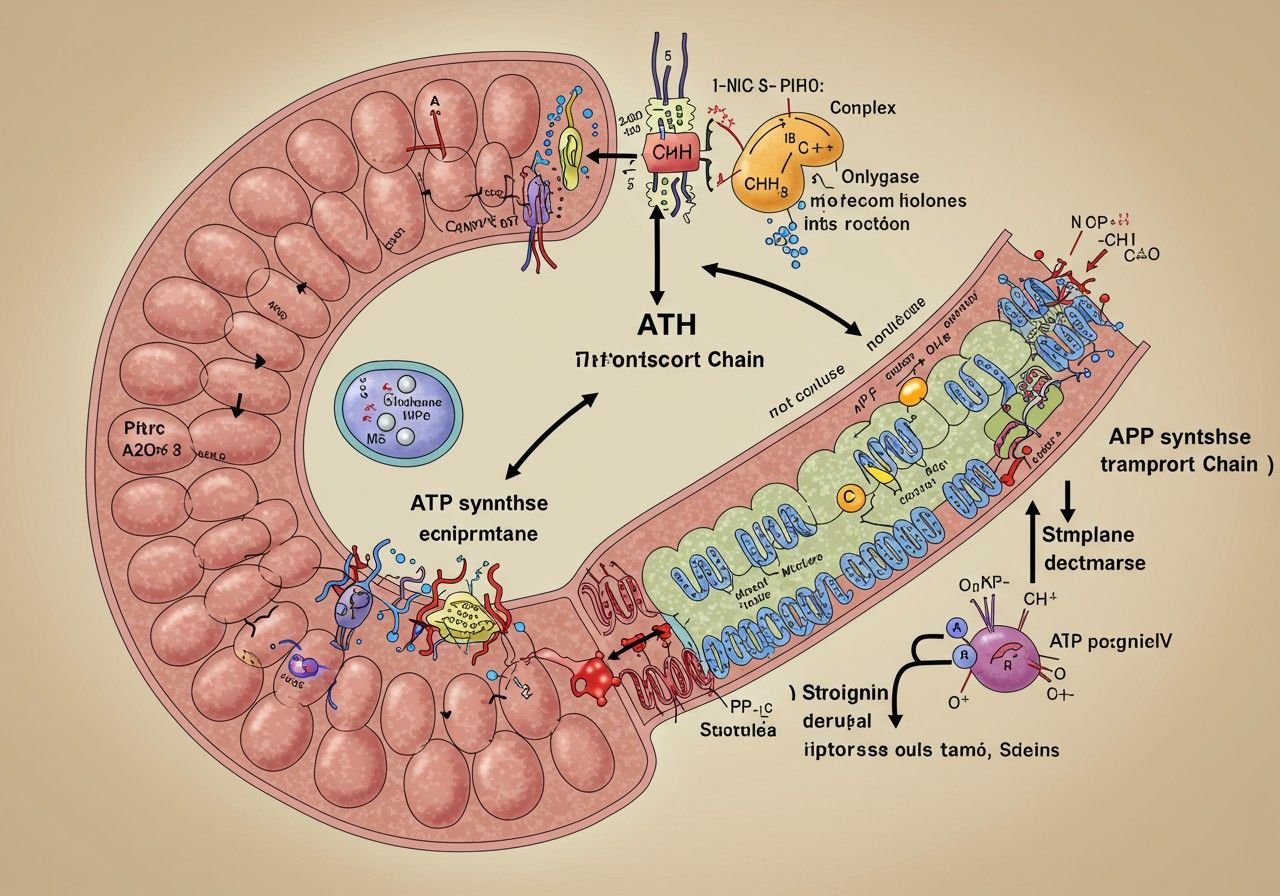
A mitokondrium, az energiagyár: a légzési lánc helyszíne
A légzési lánc működésének megértéséhez elengedhetetlen, hogy alaposan megismerjük a helyszínt, ahol ez a komplex folyamat zajlik: a mitokondriumot. A mitokondriumokat gyakran nevezik a sejtek „energiagyárainak”, és ez a megnevezés tökéletesen leírja a funkciójukat. Ezek az organellumok a legtöbb eukarióta sejtben megtalálhatók, és számuk a sejt energiaigényétől függően változhat. Például az izomsejtekben vagy a májsejtekben, amelyek rendkívül nagy energiafelhasználásúak, több ezer mitokondrium is lehet.
A mitokondriumok egyedi, kettős membránnal rendelkező szerkezete kulcsfontosságú a légzési lánc hatékony működéséhez. Két fő membránról beszélhetünk:
- Külső mitokondriális membrán: Ez a membrán viszonylag sima és áteresztő, számos porin nevű fehérjecsatornát tartalmaz, amelyek lehetővé teszik a kisebb molekulák, ionok szabad átjutását a citoplazma és a mitokondrium közötti térbe.
- Belső mitokondriális membrán: Ez a membrán sokkal összetettebb és rendkívül szelektív az áteresztőképességét tekintve. Jellemzője, hogy számos redőt, úgynevezett krisztát (cristae) alkot, amelyek jelentősen megnövelik a membrán felületét. Ez a nagy felület létfontosságú, mivel a légzési lánc összes komponense, valamint az ATP szintáz enzim is ebben a membránban található. A belső membrán áteresztőképességét speciális transzporterfehérjék szabályozzák, amelyek csak bizonyos molekulákat engednek át.
Ezen membránok között két fő tér található:
- Intermembrán tér (membránok közötti tér): Ez a külső és belső mitokondriális membrán közötti szűk tér. Ez a tér kémiai szempontból nagyon hasonlít a citoplazmához, mivel a külső membrán áteresztőképessége miatt a kisebb molekulák szabadon áramolhatnak ide. A légzési lánc során a protonok ebbe a térbe pumpálódnak.
- Mitokondriális mátrix: Ez a belső mitokondriális membrán által körülhatárolt belső tér. Itt zajlanak a Krebs-ciklus reakciói, a piruvát oxidációja, valamint a zsírsavak béta-oxidációja. A mátrixban számos enzim, koenzim és ion található, amelyek elengedhetetlenek ezekhez a folyamatokhoz.
A kriszták fontosságát nem lehet eléggé hangsúlyozni. A megnövelt felület lehetővé teszi, hogy nagy mennyiségű elektrontranszport lánc komplex és ATP szintáz molekula férjen el a belső membránban. Ez a koncentráció és a térbeli elrendezés biztosítja a légzési lánc rendkívüli hatékonyságát, lehetővé téve a sejt számára, hogy maximális mennyiségű ATP-t termeljen a rendelkezésre álló energiaforrásokból.
A mitokondrium tehát nem csupán egy hely, hanem egy aktív résztvevője az energiatermelésnek. Strukturális integritása és a membránok közötti különbségek alapvetőek a proton-gradiens kialakításához, amely az ATP szintézis hajtóereje. Bármilyen károsodás a mitokondrium szerkezetében vagy működésében súlyos következményekkel járhat a sejt energiaellátására nézve.
Az elektronhordozók szerepe: NADH és FADH2
A légzési lánc nem működhetne a NADH és FADH2 molekulák nélkül. Ezek az úgynevezett redukált koenzimek (elektronhordozók) kulcsfontosságúak, hiszen ők szállítják a nagy energiájú elektronokat a sejtlégzés korábbi szakaszaiból (glikolízis, piruvát oxidáció, Krebs-ciklus) a mitokondrium belső membránjában lévő elektrontranszport lánchoz. Gondoljunk rájuk úgy, mint apró „taxikra”, amelyek értékes rakományt, azaz elektronokat szállítanak a célállomásra, ahol azok energiája felszabadul és ATP-vé alakul.
A NADH (nikotinamid-adenin-dinukleotid) a sejtlégzés során a következő reakciókban keletkezik:
- Glikolízis (2 molekula NADH/glükóz)
- Piruvát oxidáció (2 molekula NADH/glükóz)
- Krebs-ciklus (6 molekula NADH/glükóz)
Összesen 10 molekula NADH keletkezik egy glükóz teljes lebontása során a légzési lánc előtt. A NADH egy hidrogénatomot és egy extra elektront képes felvenni, így redukált formában (NADH + H+) magas energiájú elektronokat hordoz. Amikor leadja ezeket az elektronokat a légzési láncnak, visszakerül oxidált formájába (NAD+), és újra részt vehet a metabolikus folyamatokban.
A FADH2 (flavin-adenin-dinukleotid) elsősorban a Krebs-ciklusban keletkezik, méghozzá a szukcinát szukcinát-dehidrogenáz enzim általi oxidációja során. Két FADH2 molekula képződik egy glükóz lebontása során. Hasonlóan a NADH-hoz, a FADH2 is két hidrogénatomot és két elektront szállít. Azonban van egy fontos különbség: a FADH2 elektronjai alacsonyabb energiaszinttel rendelkeznek, mint a NADH elektronjai, és a légzési láncban egy későbbi ponton lépnek be.
Az elektronok energiája a kémiai kötésekben tárolódik. Amikor a NADH és FADH2 leadják elektronjaikat a légzési lánc komplexeknek, az elektronok egy sor redoxireakción mennek keresztül, fokozatosan csökkenő energiaszintű állapotokba kerülve. Ez a fokozatos energiafelszabadulás teszi lehetővé, hogy a sejt ne egy robbanásszerűen felszabaduló energiát használjon fel, hanem kontrolláltan, lépésről lépésre gyűjtse be azt.
A NADH és FADH2 nem csupán szállítók; ők a biológiai „elektronpostások”, akik a sejtlégzés korábbi szakaszainak energiás üzeneteit továbbítják a végső energiaátalakító gépezethez, a légzési lánchoz.
Az elektronok áramlása ezen molekulákról a légzési lánc komplexekre egy „elektron-vízeséshez” hasonlítható. A NADH elektronjai a „vízesés” tetején lépnek be, míg a FADH2 elektronjai egy kicsit lejjebb, ami magyarázza a különböző energiaszintjüket és az általuk termelt ATP mennyiségének különbségét. A NADH-ról származó elektronok jellemzően több ATP-t generálnak, mint a FADH2-ről származók, mivel hosszabb utat tesznek meg a láncban, és több protonpumpálási lépést aktiválnak.
A légzési lánc komponensei: a komplexek

A légzési lánc a mitokondrium belső membránjában elhelyezkedő négy nagy fehérjekomplexből (I, II, III, IV) és két mobil elektronhordozóból (ubikinon/Koenzim Q és citokróm c) áll. Ezek a komponensek szigorú sorrendben működnek együtt, hogy az elektronok energiáját felhasználva protonokat pumpáljanak az intermembrán térbe, létrehozva ezzel az ATP szintézishez szükséges gradienset. Vizsgáljuk meg ezeket a komplexeket részletesebben.
Komplex I: NADH-dehidrogenáz
A Komplex I, más néven NADH-dehidrogenáz, az elektrontranszport lánc első és legnagyobb komplexe. Feladata, hogy fogadja a NADH által szállított elektronokat. A NADH leadja két elektronját a komplexnek, miközben visszaoxidálódik NAD+-szá. Ezek az elektronok először egy flavin-mononukleotid (FMN) kofaktorhoz, majd több vas-kén centrumhoz (Fe-S klaszterek) jutnak el a komplexen belül. Az elektronok áramlása a Komplex I-en keresztül elegendő energiát szabadít fel ahhoz, hogy négy protont (H+) pumpáljon ki a mitokondriális mátrixból az intermembrán térbe. Ez az első lépés a proton-gradiens kialakításában.
Komplex II: Szukcinát-dehidrogenáz
A Komplex II, vagy szukcinát-dehidrogenáz, egy különleges komplex, mert ez az egyetlen enzim, amely egyszerre része a Krebs-ciklusnak és a légzési láncnak. Ennek a komplexnek a feladata, hogy a FADH2 által szállított elektronokat bevezesse a láncba. A Krebs-ciklusban a szukcinát szukcinát-dehidrogenáz enzim hatására fumarátá alakul, miközben FADH2 keletkezik. A FADH2 leadja elektronjait a Komplex II-nek, amely FAD-t és vas-kén centrumokat is tartalmaz. Fontos megjegyezni, hogy a Komplex II nem pumpál protonokat az intermembrán térbe. Az általa továbbított elektronok alacsonyabb energiaszinttel rendelkeznek, mint a Komplex I-ből érkezők, és közvetlenül a következő mobil elektronhordozóhoz, az ubikinonhoz jutnak.
Ubikinon (Koenzim Q): A mobil elektronhordozó
Az ubikinon, más néven Koenzim Q (CoQ), egy lipidoldékony molekula, amely szabadon mozog a mitokondrium belső membránjában. Ez a molekula hidrofób természete miatt képes beágyazódni a membránba és híd szerepet betölteni a Komplex I/II és a Komplex III között. Az ubikinon felveszi az elektronokat a Komplex I-ből (NADH-ról származó elektronok) és a Komplex II-ből (FADH2-ről származó elektronok), majd redukált formájában (ubikinol, QH2) továbbítja azokat a Komplex III-nak. Ez a mobilitás kulcsfontosságú az elektronok hatékony átadásához a fixen rögzített komplexek között.
Komplex III: Citokróm bc1 komplex
A Komplex III, vagy citokróm bc1 komplex, a következő állomás az elektronok útján. Ez a komplex az ubikinoltól (QH2) fogadja az elektronokat. A Komplex III tartalmaz vas-kén centrumokat és kétféle citokróm b-t, valamint egy citokróm c1-et. Az elektronok áthaladása ezen a komplexen keresztül elegendő energiát szolgáltat ahhoz, hogy további négy protont pumpáljon ki a mátrixból az intermembrán térbe. A Komplex III-ban zajló elektronátadás különösen összetett, és az úgynevezett Q-ciklus mechanizmusán keresztül történik, ami biztosítja a protonpumpálás hatékonyságát és az elektronok továbbadását a következő mobil hordozónak, a citokróm c-nek.
Citokróm c: A másik mobil elektronhordozó
A citokróm c egy kis, vízoldékony fehérje, amely az intermembrán térben található. Feladata, hogy az elektronokat a Komplex III-tól a Komplex IV-hez szállítsa. Mivel vízoldékony, képes szabadon diffundálni az intermembrán térben, és a belső membrán külső felületén kötődik a komplexekhez. Minden citokróm c molekula egy elektront képes szállítani egy vasatom (hem csoport) segítségével. Ez a mobil hordozó biztosítja a gyors és hatékony elektronátadást a két nagy komplex között.
Komplex IV: Citokróm c oxidáz
A Komplex IV, más néven citokróm c oxidáz, az elektrontranszport lánc utolsó komplexe. Ez a komplex fogadja az elektronokat a citokróm c-től. A Komplex IV tartalmaz réz centrumokat (CuA, CuB) és citokróm a, valamint citokróm a3 hem csoportokat. Itt történik az elektronok végső átadása az oxigénnek. Négy elektron és négy proton egyesül egy molekula oxigénnel (O2), hogy két molekula vizet (H2O) képezzen. Ez a reakció teszi az oxigént a végső elektronakceptorrá. Az elektronok áthaladása a Komplex IV-en keresztül további két protont pumpál ki a mátrixból az intermembrán térbe. Ez a komplex kulcsfontosságú, mert az oxigén hiányában az egész légzési lánc leállna.
Összefoglalva, a négy komplex (I, III, IV) és a két mobil hordozó (ubikinon, citokróm c) egy szorosan összehangolt rendszert alkot. Az elektronok áramlása ezen a láncon keresztül fokozatosan szabadítja fel az energiát, amelyet a komplexek arra használnak fel, hogy protonokat pumpáljanak a mátrixból az intermembrán térbe. Ez a protonpumpálás hozza létre azt a elektrokémiai gradienst, amely az ATP szintézis hajtóereje.
A protonpumpa működése: elektrokémiai gradiens létrehozása
A légzési lánc lényegi működése a protonok (H+) pumpálásán alapul. Ahogy az elektronok áramlanak a Komplex I-en, III-on és IV-en keresztül, a felszabaduló energia felhasználódik arra, hogy a mitokondriális mátrixból protonokat juttassanak az intermembrán térbe. Ezt a folyamatot protonpumpálásnak nevezzük, és ez hozza létre az úgynevezett elektrokémiai gradienst, vagy más néven proton-mozgató erőt (PMF). Ez a gradiens a légzési lánc igazi „motorja”, amely végül az ATP szintézisét hajtja.
Az elektrokémiai gradiens két fő komponensből áll:
- Kémiai gradiens (pH-gradiens): Mivel a protonok a mátrixból az intermembrán térbe pumpálódnak, a mátrixban csökken a protonkoncentráció, míg az intermembrán térben növekszik. Ez azt jelenti, hogy az intermembrán tér savasabbá, a mátrix pedig lúgosabbá válik. A pH különbség (ΔpH) a kémiai gradiens.
- Elektromos potenciálkülönbség (membránpotenciál): A protonok pozitív töltésűek. Amikor pozitív töltésű részecskék elhagyják a mátrixot és felhalmozódnak az intermembrán térben, az intermembrán tér pozitívabb töltésűvé válik a mátrixhoz képest. Ez a töltéskülönbség elektromos potenciálkülönbséget (ΔΨ) hoz létre a belső mitokondriális membrán két oldala között.
Ez a két komponens, a pH-gradiens és az elektromos potenciálkülönbség együttesen alkotja a proton-mozgató erőt. Képzeljük el úgy, mint egy duzzasztógát mögött felgyűlt vizet. A víz magassága (pH-gradiens) és a benne rejlő potenciális energia (elektromos potenciálkülönbség) együttesen jelenti a felgyülemlett erőt. Ez az erő arra készteti a protonokat, hogy visszatérjenek a mátrixba, de ezt csak speciális csatornákon keresztül tehetik meg.
A légzési lánc protonpumpálása olyan, mint egy biológiai akkumulátor töltése: az elektronok energiája egy potenciális energiaforrást hoz létre, amely később az ATP szintézisét hajtja.
A protonok pumpálásának mechanizmusa a komplexeken belül összetett, és magában foglalja a fehérjék konformációs változásait, amelyek a protonok átjutását teszik lehetővé. A Komplex I, III és IV különböző számú protont pumpál:
- Komplex I: 4 proton / NADH elektronpár
- Komplex III: 4 proton / elektronpár (Q-ciklus révén)
- Komplex IV: 2 proton / elektronpár
A Komplex II, mint már említettük, nem pumpál protonokat. Ez az oka annak, hogy a FADH2-ről származó elektronok kevesebb ATP-t termelnek, mint a NADH-ról származók, mivel az FADH2 elektronjai „kihagyják” a Komplex I protonpumpáló lépését.
A proton-mozgató erő fenntartása kritikus fontosságú. Ha a belső mitokondriális membrán károsodik és áteresztővé válik a protonok számára (például bizonyos méreganyagok hatására), a gradiens összeomlik, és az ATP termelés leáll, még akkor is, ha az elektronok tovább áramlanak a láncban. Ez a jelenség az szétkapcsolás, amely rendkívül veszélyes lehet a sejtre nézve.
Az elektrokémiai gradiens létrehozása egy elegáns és rendkívül hatékony módja az energia tárolásának. Ez a gradiens nemcsak az ATP szintéziséhez szükséges energiát biztosítja, hanem más fontos folyamatokat is hajt a mitokondriumban, például az anyagcsere-köztes termékek vagy ionok transzportját a membránon keresztül. Ez a központi szerep teszi a proton-mozgató erőt az egyik legfontosabb energetikai alapelvvé a biológiai rendszerekben.
Az ATP szintézis: az ATP szintáz csodája
Miután a légzési lánc komplexek sikeresen létrehozták az elektrokémiai gradienst, a felhalmozott potenciális energiát át kell alakítani a sejtek számára hasznosítható ATP-vé. Ez a feladat az ATP szintáz nevű rendkívüli enzimre hárul. Az ATP szintáz a mitokondrium belső membránjában található, és egy molekuláris „turbinához” hasonlít, amely a protonok áramlását használja fel az ATP előállítására. Ez a folyamat az oxidatív foszforiláció néven ismert, mivel az ATP szintézise az oxidatív folyamatok (elektrontranszport) energiájával párosul.
Az ATP szintáz két fő részből áll:
- F0 egység: Ez a rész a belső mitokondriális membránba ágyazódik be, és egy protontranszport csatornát alkot. A protonok ezen a csatornán keresztül áramlanak vissza az intermembrán térből a mátrixba, a koncentráció- és töltésgradiens mentén. Az F0 egység egy gyűrű alakú c-alegységből és egy a-alegységből áll, amelyek a protonok áramlását szabályozzák.
- F1 egység: Ez a rész a mitokondriális mátrixba nyúlik be, és itt zajlik az ATP tényleges szintézise. Az F1 egység hat alegységből (három α és három β alegység) áll, amelyek egy központi γ-tengely körül helyezkednek el. A β alegységek tartalmazzák az aktív centrumokat, ahol az ADP és a szervetlen foszfát (Pi) ATP-vé alakul.
A működési elv a következő: Ahogy a protonok az F0 egységen keresztül áramlanak vissza a mátrixba, az F0 egység c-gyűrűje forogni kezd. Ez a forgás mechanikai energiát generál, amelyet a γ-tengely továbbít az F1 egységhez. A γ-tengely forgása konformációs változásokat idéz elő az F1 egység β alegységeiben. Ez az úgynevezett kötésváltozási mechanizmus, amelyet Paul Boyer írt le, és amiért Nobel-díjat kapott.
A β alegységek három különböző konformációs állapotot vesznek fel rotáció közben:
- L (Loose) állapot: Lazán köti az ADP-t és a Pi-t.
- T (Tight) állapot: Szorosan köti az ADP-t és a Pi-t, és katalizálja az ATP képződését.
- O (Open) állapot: Alacsony affinitással rendelkezik az ATP-hez, így az újonnan szintetizált ATP felszabadul.
Ahogy a γ-tengely forog, a β alegységek ciklikusan váltogatják ezeket az állapotokat, folyamatosan kötve az ADP-t és Pi-t, szintetizálva az ATP-t, majd felszabadítva azt. Körülbelül 3-4 proton áthaladása szükséges egy ATP molekula szintéziséhez.
Az ATP szintáz egy molekuláris csoda, amely a protonok áramlásának energiáját egy forgó mechanizmuson keresztül alakítja át kémiai energiává, létrehozva a sejt univerzális valutáját, az ATP-t.
Az oxidatív foszforiláció a légzési lánc és az ATP szintáz együttes működésének eredménye. Ez a folyamat rendkívül hatékony. Egy glükóz molekula teljes aerob lebontása során akár 30-32 ATP molekula is keletkezhet, szemben a glikolízis során keletkező mindössze 2 ATP-vel. Ez a hatalmas energiahozam teszi az oxidatív foszforilációt a legtöbb aerob élőlény energiaellátásának alapkövévé.
Az ATP szintáz működése rendkívül finoman szabályozott, hogy a sejt energiaigényéhez igazodjon. Az ATP/ADP arány például közvetlenül befolyásolja az enzim aktivitását. Ha sok az ATP, az enzim gátolt, ha sok az ADP, az enzim aktívabbá válik, jelezve, hogy a sejtnek több energiára van szüksége. Ez a szabályozás biztosítja, hogy a sejt ne termeljen feleslegesen energiát, és ne pazarolja el erőforrásait.
Az oxigén szerepe: a végső elektronakceptor
A légzési lánc működésének egyik alapvető feltétele az oxigén jelenléte. Az oxigénnek kulcsfontosságú szerepe van, hiszen ő a végső elektronakceptor. Ez azt jelenti, hogy az elektrontranszport lánc végén, a Komplex IV-nél az oxigén fogadja a láncban végigszállított elektronokat. E nélkül az utolsó lépés nélkül az egész rendszer leállna, és az ATP termelés drámaian csökkenne.
A Komplex IV-ben négy elektron és négy proton (a mitokondriális mátrixból) egyesül egy molekula oxigénnel (O2), hogy két molekula vizet (H2O) képezzen:
O2 + 4e- + 4H+ → 2H2O
Ez a reakció létfontosságú több okból is:
- Az elektronok áramlásának fenntartása: Az oxigén extrém módon elektronegatív, ami azt jelenti, hogy erős vonzást gyakorol az elektronokra. Ez a vonzás „húzza” az elektronokat végig a légzési láncon. Ha nincs oxigén, az elektronoknak nincs hová menniük a Komplex IV-től, így feltorlódnak a láncban. Ez visszahat a korábbi komplexekre, megakadályozva, hogy azok felvegyék az elektronokat a NADH-tól és FADH2-től. Ennek eredményeként a NADH és FADH2 nem tud visszaoxidálódni NAD+-ra és FAD-ra.
- A NAD+ és FAD regenerálása: A sejtlégzés korábbi szakaszai (glikolízis, Krebs-ciklus) csak akkor tudnak tovább működni, ha a NADH és FADH2 visszaoxidálódik NAD+-ra és FAD-ra. Az oxigén hiányában ezek a koenzimek redukált formában maradnak, és a Krebs-ciklus, valamint a glikolízis is leáll (vagy alternatív útvonalra terelődik, mint az anaerob légzés).
- Az aerob élet alapja: Az oxigén mint végső elektronakceptor teszi lehetővé az aerob anyagcserét, amely sokkal hatékonyabb az ATP termelésben, mint az anaerob folyamatok. Ez az oka annak, hogy a legtöbb komplex élőlény, beleértve az embert is, oxigénre van utalva a túléléshez.
Mi történik, ha nincs elegendő oxigén? Ez az állapot az anoxia (teljes oxigénhiány) vagy hipoxia (részleges oxigénhiány). Ilyen körülmények között a légzési lánc leáll. A sejt kénytelen alternatív, sokkal kevésbé hatékony energiaforrásokhoz nyúlni. Az emberi testben ez a tejsavas erjedés. A glikolízis továbbra is termel 2 ATP-t, de a piruvát nem lép be a mitokondriumba. Ehelyett a piruvát tejsavvá alakul, miközben a NADH visszaoxidálódik NAD+-ra, így a glikolízis folytatódhat. Ez azonban csak rövid távon fenntartható megoldás, és a felgyülemlett tejsav izomfájdalmat és fáradtságot okozhat.
Az oxigén nem csupán a levegő alkotóeleme; a légzési láncban betöltött szerepe révén ő a láthatatlan motor, amely az élet energiafolyamatainak végső lökését adja.
Az oxigén tehát nem csupán passzív résztvevő; aktívan „húzza” az elektronokat a láncban, lehetővé téve a folyamatos ATP termelést. Az oxigén hiánya vagy elégtelen mennyisége azonnali és súlyos következményekkel jár a sejtek energiaellátására nézve, ami magyarázza, miért olyan kritikus az oxigénellátás a szervezet számára.
A légzési lánc szabályozása

A sejtek energiaigénye folyamatosan változik, így a légzési lánc működését is szigorúan szabályozni kell, hogy a sejt ne termeljen feleslegesen energiát, és ne pazarolja el erőforrásait. A szabályozás elsősorban a rendelkezésre álló szubsztrátok (NADH, FADH2), az ADP/ATP arány és az oxigénkoncentráció függvényében történik.
A rendelkezésre álló szubsztrátok
Az elektronhordozó molekulák, a NADH és FADH2, a légzési lánc „üzemanyagai”. Ha ezekből kevés van, az elektrontranszport lánc lelassul. Ezen molekulák termelődését a glikolízis és a Krebs-ciklus szabályozza, amelyek viszont a sejt energiaállapotától és a táplálékforrások elérhetőségétől függenek. Például, ha sok a glükóz, a glikolízis aktívabb, több NADH keletkezik, ami felpörgeti a légzési láncot.
Az ATP/ADP arány
Az ATP/ADP arány az egyik legfontosabb szabályozó tényező, amely közvetlenül jelzi a sejt energiaállapotát.
- Ha az ATP szint magas és az ADP szint alacsony, az azt jelzi, hogy a sejtnek elegendő energiája van. Ebben az esetben a légzési lánc és az ATP szintáz aktivitása csökken. Az ATP például alloszterikusan gátolja a Komplex IV-et, lassítva az elektronáramlást.
- Ha az ADP szint magas és az ATP szint alacsony, az a sejt energiahiányát jelzi. Ekkor az ADP aktiválja az ATP szintázt, és növeli a légzési lánc komplexek aktivitását, felgyorsítva az ATP termelést. Ez a szoros kapcsolat biztosítja, hogy a sejt pontosan annyi ATP-t termeljen, amennyire éppen szüksége van. Ezt a jelenséget respirációs kontrollnak nevezik.
Oxigénkoncentráció
Ahogy már említettük, az oxigén a végső elektronakceptor. Ha az oxigénkoncentráció alacsony, a légzési lánc lelassul vagy leáll, függetlenül attól, hogy mennyi NADH és FADH2 áll rendelkezésre. Ez az alapja az anaerob anyagcsere bekapcsolásának oxigénhiányos állapotokban.
Szétkapcsoló fehérjék (UCP-k)
Léteznek speciális fehérjék, az úgynevezett szétkapcsoló fehérjék (Uncoupling Proteins, UCPs), amelyek képesek „szétkapcsolni” az elektrontranszportot az ATP szintézisétől. Ezek a fehérjék protocsatornákat hoznak létre a belső mitokondriális membránban, lehetővé téve a protonok számára, hogy visszaszivárogjanak a mátrixba anélkül, hogy az ATP szintázon keresztül haladnának. Amikor a protonok az UCP-n keresztül jutnak vissza, a gradiensben tárolt energia hő formájában szabadul fel, ATP helyett. Ez a mechanizmus különösen fontos az újszülöttek barna zsírszövetében, ahol a termogenezis (hőtermelés) célját szolgálja.
A légzési lánc szabályozása egy kifinomult tánc, ahol a sejt energiamérlege határozza meg a ritmust, biztosítva az optimális ATP-termelést a változó igényekhez igazodva.
Hormonális szabályozás
Bizonyos hormonok, például a pajzsmirigyhormonok, befolyásolhatják a mitokondriumok számát és aktivitását, ezáltal közvetetten hatva a légzési lánc kapacitására. A pajzsmirigyhormonok növelhetik az UCP-k expresszióját is, fokozva a hőtermelést és az alapanyagcserét.
A légzési lánc komplex szabályozása biztosítja, hogy a sejt energiaellátása mindig optimális legyen, elkerülve az energiahiányt vagy a felesleges energiatermelést, ami mindkettő káros lehet a sejt számára.
A légzési lánc zavarai és betegségek
A légzési lánc rendkívül összetett és precízen működő gépezet, amelynek zavarai súlyos következményekkel járhatnak a sejtek, szövetek és az egész szervezet számára. Mivel az ATP termelés nagyrészt ezen a folyamaton keresztül zajlik, bármilyen hiba az elektrontranszport láncban vagy az ATP szintázban drámaian befolyásolhatja a sejt energiaellátását, és számos betegség kialakulásához vezethet.
Mérgező anyagok hatása
Számos vegyület képes gátolni a légzési lánc különböző pontjait, ezzel megzavarva az ATP termelést:
- Cianid: Ez a rendkívül mérgező anyag a Komplex IV-hez kötődik, és irreverzibilisen gátolja annak működését. A Komplex IV leállásával az elektronok nem tudnak tovább áramlani az oxigénhez, feltorlódnak az egész láncban, és az ATP termelés leáll. Ez magyarázza a cianid gyors és halálos hatását.
- Szén-monoxid (CO): Hasonlóan a cianidhoz, a szén-monoxid is a Komplex IV-hez kötődik, de kevésbé szorosan. Verseng az oxigénnel a kötőhelyért, és gátolja a Komplex IV működését, ami oxigénhiányos állapothoz hasonló tüneteket okoz a sejtekben.
- Rotenon és amital: Ezek az anyagok a Komplex I-et gátolják, megakadályozva a NADH-ról származó elektronok belépését a láncba.
- Antimicin A: Ez a vegyület a Komplex III-at gátolja.
- Oligomicin: Ez az antibiotikum az ATP szintáz F0 alegységéhez kötődik, gátolva a protonok áramlását és az ATP szintézisét.
Ezek a mérgek rávilágítanak a légzési lánc sérülékenységére és arra, hogy mennyire alapvető a hibátlan működése az élethez.
Mitokondriális betegségek
A mitokondriális betegségek egy heterogén csoportja olyan genetikai rendellenességeknek, amelyek a mitokondriumok működését érintik. Ezek a betegségek érinthetik a légzési lánc bármelyik komplexét, az ATP szintázt, vagy a mitokondriális DNS-t (mtDNS) is. Mivel a mitokondriumok minden sejtben jelen vannak, a betegségek tünetei rendkívül sokfélék lehetnek, de leggyakrabban azokat a szerveket érintik, amelyeknek nagy az energiaigénye:
- Idegrendszer: Epilepszia, stroke-szerű epizódok, demencia, ataxia (mozgáskoordinációs zavarok).
- Izomrendszer: Izomgyengeség, izomfájdalom, izomsorvadás (miopátia).
- Szív: Kardiomiopátia (szívizombetegség).
- Máj: Májműködési zavarok.
- Vese: Veseelégtelenség.
- Endokrin rendszer: Cukorbetegség, pajzsmirigy alulműködés.
Ezek a betegségek örökölhetők anyai ágon (mivel a mitokondriumok a petesejtből származnak) vagy autoszomális recesszív/domináns módon. A diagnózis és a kezelés rendkívül nehéz, és gyakran tüneti kezelésre korlátozódik.
A légzési lánc zavarai nem csupán elméleti problémák; valós, pusztító betegségeket okozhatnak, rávilágítva arra, hogy az élet energetikai alapjai mennyire törékenyek lehetnek.
A reaktív oxigénfajták (ROS) és az oxidatív stressz
A légzési lánc működése során, különösen a Komplex I és Komplex III-ban, előfordulhat, hogy az elektronok nem jutnak el az oxigénhez, hanem részlegesen redukálják azt, reaktív oxigénfajtákat (ROS) hozva létre. Ilyenek például a szuperoxid anion (O2•-) és a hidrogén-peroxid (H2O2). Ezek a ROS rendkívül reaktív molekulák, amelyek károsíthatják a sejtek makromolekuláit (DNS, fehérjék, lipidek), ami oxidatív stresszhez vezet. Az oxidatív stressz hozzájárul az öregedési folyamatokhoz és számos krónikus betegség, például a rák, a neurodegeneratív betegségek és a szív- és érrendszeri betegségek kialakulásához.
A sejt rendelkezik antioxidáns védekező rendszerekkel (pl. szuperoxid-diszmutáz, kataláz, glutation-peroxidáz), amelyek semlegesítik a ROS-t. Azonban, ha a ROS termelése meghaladja a védekező kapacitást, károsodás lép fel. A légzési lánc hatékony és kontrollált működése tehát nemcsak az ATP termelés, hanem a sejt integritásának fenntartása szempontjából is kritikus.
A légzési lánc evolúciója és jelentősége
A légzési lánc egyike a biológia legősibb és legfontosabb anyagcsere-útjainak. Jelentősége nem csupán az egyedi sejt működésében rejlik, hanem abban is, hogy alapvető szerepet játszott az élet evolúciójában, különösen a komplex, többsejtű szervezetek kialakulásában. Az aerob légzés megjelenése, amelynek a légzési lánc a központi eleme, óriási előnyt jelentett az anaerob életformákkal szemben.
Az evolúciós eredet
A légzési lánc komponenseinek eredete valószínűleg a korai prokarióta (baktériumok és archeák) sejtekig vezethető vissza, amelyek már milliárd évekkel ezelőtt használtak elektrontranszport láncokat az energia előállítására. Ezek a korai láncok valószínűleg kevésbé voltak hatékonyak, és különböző elektronakceptorokat használhattak az oxigén helyett (pl. szulfát, nitrát). Amikor a Föld légköre oxigénben gazdagabbá vált a fotoszintetizáló szervezetek elterjedésével, az oxigén mint végső elektronakceptor használata hatalmas evolúciós nyomást gyakorolt. Az oxigén magas elektronegatívitása lehetővé tette a sokkal hatékonyabb energiafelszabadítást, ami az ATP szintézis robbanásszerű növekedését eredményezte.
A mitokondriumok, mint organellumok, az endoszimbiózis elmélete szerint egy ősi prokarióta sejt (valószínűleg egy aerob baktérium) bekebelezésével jöttek létre egy nagyobb, ősi eukarióta sejt által. Ez a szimbiotikus kapcsolat kölcsönösen előnyös volt: a bekebelezett baktérium (a későbbi mitokondrium) hatékony ATP-termelést biztosított a gazdasejt számára, míg a gazdasejt stabil környezetet és táplálékot nyújtott. Ez a „fúzió” alapozta meg a komplex eukarióta életformák, így az állatok, növények és gombák kialakulását és fejlődését.
Az aerob életforma alapja
Az aerob légzés, a légzési lánccal a középpontjában, sokkal nagyobb mennyiségű ATP-t termel egy glükóz molekulából, mint az anaerob folyamatok. Ez a megnövelt energiahozam tette lehetővé a nagyobb testméret, a komplexebb sejtszerkezetek és a nagyobb energiaigényű életfolyamatok kialakulását. Gondoljunk csak az agy működésére, az izmok összehúzódására vagy a testhőmérséklet fenntartására – mindezek hatalmas mennyiségű ATP-t igényelnek, amit az anaerob anyagcsere nem tudna biztosítani.
A légzési lánc nem csupán egy biokémiai útvonal; az élet motorja, amely az ősi Földön lehetővé tette a komplex életformák kialakulását, és ma is minden aerob élőlény létezésének alapját képezi.
A légzési lánc jelentősége napjainkban
A légzési lánc működése nemcsak az evolúcióban volt kulcsfontosságú, hanem a modern biológia és orvostudomány számára is kiemelten fontos terület. A légzési lánc zavarai számos betegség gyökerét képezik, a neurodegeneratív kórképektől (pl. Parkinson-kór, Alzheimer-kór) a metabolikus szindrómáig és a rákig. A légzési lánc kutatása segíthet új terápiás stratégiák kidolgozásában ezekre a betegségekre, például mitokondrium-specifikus gyógyszerek vagy antioxidánsok fejlesztésében.
Ezenkívül a légzési lánc a szabadgyökök termelődésének egyik fő helye is, ami az oxidatív stressz és az öregedés folyamataiban játszik szerepet. Ennek megértése alapvető fontosságú az öregedés elleni stratégiák és az egészségmegőrzés szempontjából. A légzési lánc tehát egy olyan alapvető biokémiai folyamat, amely nélkülözhetetlen az élethez, és amelynek mélyebb megértése kulcsot adhat számos egészségügyi kihívás megoldásához.
Összefüggések más anyagcsere-utakkal
A légzési lánc nem egy elszigetelt biokémiai útvonal, hanem szorosan integrálódik a sejt teljes anyagcsere-hálózatába. Kapcsolata a glikolízissel és a Krebs-ciklussal már világos, de jelentős interakciói vannak a lipid- és fehérjeanyagcserével is, biztosítva a sejt számára a rugalmas energiaellátást és a makromolekulák szintéziséhez szükséges prekurzorokat.
Szénhidrát-anyagcsere
A glikolízis és a Krebs-ciklus a szénhidrátok (elsősorban glükóz) lebontásának fő útvonalai. Ezek a folyamatok termelik a légzési lánc számára szükséges NADH és FADH2 molekulákat. A glükóz tehát a légzési lánc egyik fő „üzemanyaga”. A glikolízis során keletkező piruvát acetil-CoA-vá alakul, amely belép a Krebs-ciklusba, ezzel táplálva a légzési láncot. Ez a szoros kapcsolat biztosítja, hogy a szénhidrátokból származó energia maximális hatékonysággal hasznosuljon.
Lipid-anyagcsere
A zsírok, vagy lipidek, a szervezet legfontosabb energiaraktárai. A zsírsavak lebontása, az úgynevezett béta-oxidáció, a mitokondriális mátrixban zajlik. Ennek során acetil-CoA, NADH és FADH2 keletkezik. Az acetil-CoA belép a Krebs-ciklusba, míg a NADH és FADH2 közvetlenül a légzési láncba szállítja elektronjait. Ez azt jelenti, hogy a zsírok lebontásából származó energia is a légzési láncon keresztül alakul ATP-vé. A zsírsavak jóval több ATP-t termelnek, mint a glükóz, mivel hosszabb szénláncuk révén több acetil-CoA-t, NADH-t és FADH2-t generálnak.
Például egy 16 szénatomos palmitinsav molekula teljes lebontása során körülbelül 106 ATP molekula keletkezik, szemben a glükóz 30-32 ATP-jével. Ez a magyarázata annak, hogy a zsírok miért jelentenek hatékonyabb energiaforrást.
Fehérje-anyagcsere
A fehérjék lebontása során keletkező aminosavak is felhasználhatók energiatermelésre, különösen éhezés vagy szénhidrátok hiánya esetén. Az aminosavak deaminálódás után különböző metabolikus intermedierekké alakulhatnak, amelyek beléphetnek a glikolízisbe, a piruvát oxidációba vagy közvetlenül a Krebs-ciklusba. Ezek az intermedierek, mint például a piruvát, acetil-CoA, oxálacetát, szukcinil-CoA vagy α-ketoglutarát, tovább oxidálódnak, és NADH, valamint FADH2 termelődéséhez vezetnek, amelyek aztán a légzési láncba juttatják elektronjaikat.
A fehérje-anyagcsere és a légzési lánc közötti kapcsolat biztosítja, hogy a szervezet extrém körülmények között is képes legyen energiát termelni, még akkor is, ha a szénhidrát- és zsírraktárak kimerültek.
A légzési lánc a sejt metabolikus hálózatának központi csomópontja, amely rugalmasan képes felhasználni a szénhidrátokból, zsírokból és fehérjékből származó energiát, biztosítva a folyamatos ATP-ellátást minden életfolyamathoz.
A légzési lánc és a metabolikus rugalmasság
A légzési lánc központi szerepe a különböző makromolekulák lebontásában a sejt metabolikus rugalmasságának alapját képezi. Ez a rugalmasság lehetővé teszi a sejt számára, hogy alkalmazkodjon a változó táplálékforrásokhoz és energiaigényekhez. Akár glükóz, akár zsírsavak, akár aminosavak állnak rendelkezésre, a légzési lánc képes felhasználni az azokból származó elektronokat az ATP hatékony előállítására. Ez a adaptációs képesség kulcsfontosságú az élőlények túléléséhez a változó környezeti feltételek között.
Ez a mély integráció azt is jelenti, hogy a légzési lánc zavarai nem csak közvetlenül az ATP-termelést érintik, hanem visszahatnak a teljes anyagcserére, befolyásolva a szénhidrát-, zsír- és fehérje-anyagcserét is. Ennek megértése kulcsfontosságú a metabolikus betegségek patomechanizmusának feltárásában és új terápiás célpontok azonosításában.
Gyakori tévhitek és tisztázások a légzési lánccal kapcsolatban
A légzési lánc bonyolultsága miatt számos tévhit és félreértés kering a köztudatban, különösen a laikusok körében. Fontos tisztázni ezeket, hogy a folyamat megértése pontos és tudományosan megalapozott legyen.
Tévhit 1: A légzési lánc „égeti” a táplálékot.
Tisztázás: Bár a légzési lánc oxidatív folyamat, és energiafelszabadulással jár, nem szó szerint „égeti” a táplálékot. Az égés egy gyors, kontrollálatlan oxidációs folyamat, amely nagy mennyiségű hőt szabadít fel egyszerre. A légzési láncban az oxidáció egy sor apró, kontrollált lépésben zajlik, ahol az energia fokozatosan szabadul fel, és nem hő, hanem ATP formájában tárolódik. Ez a finomhangolt, lépcsőzetes folyamat sokkal hatékonyabb az energia kinyerésében.
Tévhit 2: Az oxigén közvetlenül ATP-t termel.
Tisztázás: Az oxigén a végső elektronakceptor a légzési láncban. Nélküle az egész rendszer leállna, de az oxigén önmagában nem termel ATP-t. Az ATP-t az ATP szintáz enzim állítja elő, felhasználva a proton-mozgató erőt, amelyet a légzési lánc komplexek hoztak létre a protonok pumpálásával. Az oxigén szerepe az elektronok elvezetésében és a lánc folyamatos működésének biztosításában rejlik.
Tévhit 3: A légzési lánc csak a glükóz lebontásából származó energiát hasznosítja.
Tisztázás: Bár a glükóz a sejtek elsődleges energiaforrása, a légzési lánc rendkívül rugalmas. Amint azt már tárgyaltuk, képes felhasználni a zsírsavak és az aminosavak lebontásából származó elektronokat is. A NADH és FADH2 molekulák a glükóz mellett a zsír- és fehérjeanyagcsere során is keletkeznek, és ezek is bekerülnek az elektrontranszport láncba, ahol az energiájuk ATP-vé alakul.
Tévhit 4: A légzési lánc az egyetlen ATP-termelő folyamat a sejtben.
Tisztázás: A légzési lánc az aerob ATP-termelés legnagyobb részéért felelős, de nem az egyetlen. A glikolízis közvetlenül 2 ATP-t termel szubsztrát szintű foszforilációval, és a Krebs-ciklus is termel 1 GTP-t (ami könnyen ATP-vé alakul) ciklusonként. Ezek a mennyiségek eltörpülnek a légzési lánc által termelt ATP mellett, de fontosak, különösen oxigénhiányos állapotokban vagy a gyors energiaigény kielégítésekor.
Tévhit 5: Az egész légzési lánc egyetlen nagy enzim.
Tisztázás: A légzési lánc nem egyetlen enzim, hanem négy nagy fehérjekomplexből (I, II, III, IV) és két mobil elektronhordozóból (ubikinon és citokróm c) áll. Ezek a komponensek egymással szorosan együttműködve, de különálló egységekként működnek, mindegyiknek megvan a maga specifikus feladata az elektronok átadásában és a protonok pumpálásában.
A légzési lánc működésének alapos megértése elengedhetetlen a biokémia, az orvostudomány és az élettudományok számos területén, segítve a valós biológiai folyamatok pontos értelmezését.
Tévhit 6: Minden energiát ATP-vé alakít.
Tisztázás: Bár a légzési lánc rendkívül hatékony, nem 100%-os hatásfokkal alakítja át az energiát ATP-vé. Az energia egy része mindig hő formájában szabadul fel. Ez a hőtermelés létfontosságú a testhőmérséklet fenntartásához (különösen a melegvérű állatoknál). A szétkapcsoló fehérjék (UCP-k) például szándékosan növelik a hőtermelést az ATP-termelés rovására, ami a termogenezisben játszik szerepet.
Ezen tévhitek tisztázása segít abban, hogy a légzési lánc valós jelentőségét és működési elvét pontosan érthessük, elkerülve a leegyszerűsítéseket, amelyek torzítják a tudományos valóságot.







